

INTRODUCTION
MATERIALS AND METHODS
Breeding scheme and generation advancement
Rapid generation advancement (RGA) system
Foreground and background selection
DNA extraction and marker analysis
Phenotypic evaluation and selection of advanced lines
Evaluation of RSV resistance
Statistical analysis
RESULTS
Development and segregation analysis of RSV-resistant lines
Recurrent parent genome recovery and background selection
Agronomic traits and correlation analysis
Development and characterization of the RSV-resistant NIL ‘Milyang397’
DISCUSSION
CONCLUSION
INTRODUCTION
Rice stripe virus (RSV) is one of the most destructive viral diseases of rice, causing substantial yield losses in regions where japonica rice is predominantly grown, including China, Japan, and Korea. The virus is transmitted by the small brown planthopper (Laodelphax striatellus) through a persistent, circulative–propagative transmission mode (Cho et al., 2013). RSV was first detected in Korea in 1965 and remained largely restricted to double-cropping areas in the southern region until the 1980s. As temperatures on the Korean Peninsula increased, the disease gradually spread northward. Major outbreaks were recorded in 2001, affecting about 4,600 ha, and in 2007, when roughly 5,000 ha were damaged in the western coastal region. Subsequent surveys also reported infections in mid-mountain and highland areas, indicating that RSV can occur across the entire country (Lee et al., 2008).
Because effective control options for RSV are limited, management has largely depended on insecticides targeting its vector, the small brown planthopper (SBPH). However, intensive insecticide use has caused substantial environmental impacts. As a result, breeding RSV-resistant rice cultivars is regarded as a cost-effective and environmentally sustainable approach for long-term disease management (Hu et al., 2020). RSV is transmitted by the SBPH in a persistent, circulative–propagative mode, and its distribution is expected to shift northward as the vector’s overwintering range expands with climate warming (Matsukura et al., 2023; Yamamura & Yokozawa, 2002). Most resistant cultivars carry a single major resistance gene, such as Stvb-i, which makes them vulnerable to resistance breakdown as the virus evolves (Hayano-Saito et al., 2000; McDonald &Linde, 2002). Some of these cultivars have also shown reduced yield potential. To address this, combining marker-assisted selection (MAS) with rapid generation advance (RGA) has been proposed as an effective precision-breeding approach for developing RSV-resistant rice cultivars (Collard & Mackill, 2008).
Phenotypic analysis is a core element of crop improvement. As breeders work to increase crop productivity and ensure future food security, the need for accurate, large-scale phenotypic data continues to grow. However, conventional phenotyping, which depends on manual measurements, is labor-intensive, costly, and susceptible to human error (Atefi et al., 2021).
In contrast, molecular marker–based screening is regarded as the most efficient approach because it is not influenced by environmental variation that affects biological assays (Hasan et al., 2021). To develop molecular markers for RSV resistance, the ST-10 marker linked to the Stvb-i gene derived from the resistant variety Modan was developed (Hayano-Saito et al., 1998). However, ST-10 is applicable only to cultivars carrying the Modan-derived resistance allele and cannot distinguish resistant genotypes derived from indica or Tongil-type varieties.
During the 1960s, the Stvb-i gene from Modan was introgressed into the japonica variety Norin 8, and subsequently utilized in the breeding of RSV-resistant japonica rice cultivars in Korea and Japan. In a previous study, we developed an InDel marker closely linked to the Stvb-i gene derived from indica rice for use in marker-assisted screening (Kwon et al., 2012)
Haedamssal is an early-maturing japonica cultivar that carries Stvb-i-mediated RSV resistance, whereas Odae is an early- maturing cultivar with large grain size but susceptible to RSV. In this study, we present a rapid and effective backcross breeding strategy for developing RSV-resistant near-isogenic lines (NILs) in the Odae background by combining marker-assisted backcrossing (MABC) with a RGA system.
MATERIALS AND METHODS
Breeding scheme and generation advancement
In this study, rice lines with resistance to RSV were developed using a MABC strategy, in which Odae was used as the recurrent parent and Haedamssal as the donor parent. The initial cross was performed in the summer of 2017 to produce F1 seeds. The BC1F1 generation was obtained during the 2017/2018 winter greenhouse season, followed by BC2F1 production in the summer of 2018. The BC2F2 population was grown in the summer of 2019, and subsequent generations were advanced by one generation per year under summer field conditions until 2025.
Rapid generation advancement (RGA) system
Generation advancement was conducted in the winter greenhouse of the Department of Upland Crop Breeding, National Institute of Crop Science (NICS), Rural Development Administration (RDA), Miryang, Korea (20 Jompiljae-ro, Miryang), using a RGA system (Fig. 1), which enabled up to two generations per year. The greenhouse was maintained under natural light conditions without supplemental lighting, and the nighttime temperature was controlled at 22°C. All seeds used in this study were propagated at the Department of Upland Crop Breeding, NICS.
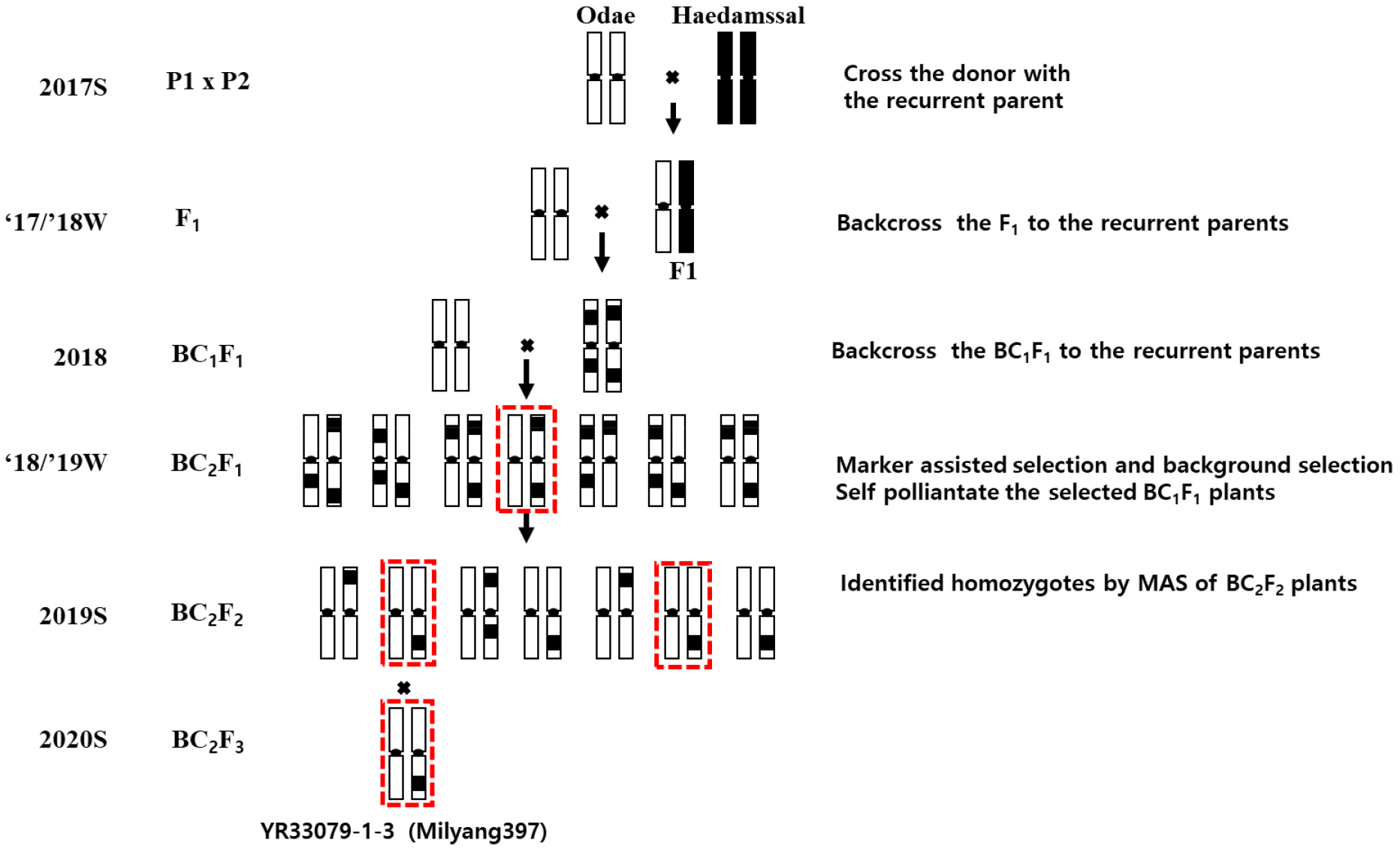
Fig. 1
Breeding scheme for developing the rice stripe virus (RSV)-resistant NIL ‘Milyang397’. The resistant donor parent ‘Haedamssal’ was crossed with the susceptible recurrent parent ‘Odae’, followed by marker-assisted backcrossing and selection using the RSV resistance-linked marker InDel7 (red boxes). The final homozygous line with high recurrent parent genome recovery was designated as ‘Milyang397’.
Foreground and background selection
For foreground selection, the RSV resistance gene was screened using a gene-specific molecular marker, InDel7. Seven homozygous and three heterozygous BC1F1 plants (Aa) carrying the target allele were selected and backcrossed to Odae to generate BC2F1 seeds. In the BC2F1 and BC2F2 generations, the same marker was used to select heterozygous (BC2F1) and homozygous (BC2F2) plants at the RSV resistance locus. To evaluate the recovery rate of the recurrent parent genome (RPG), the first background selection was performed in the BC2F1 generation using 116 polymorphic Kompetitive Allele-Specific PCR (KASP) markers, and the selected plants were self-pollinated to produce BC2F2 seeds. A second round of background selection was then carried out to maximize RPG recovery.
DNA extraction and marker analysis
Genomic DNA was extracted from fresh leaves of four-week-old seedlings using the cetyltrimethyl ammonium bromide (CTAB) method. DNA concentration and purity were measured using a NanoDrop spectrophotometer (ND-1000, Thermo Fisher Scientific, USA). The extracted DNA was diluted with 1× TE buffer (10 mM Tris-HCl, pH 8.0; 1 mM EDTA, pH 8.0) to a final concentration of 5 ng/μL and stored at 4°C for subsequent PCR analysis. Each PCR reaction was carried out in a total volume of 15 μL containing 25 ng of template DNA, 1.0 μM of each primer, 0.8 μL of dNTPs, 1.5 μL of 10× PCR buffer, 6.6 μL of double-distilled water (DDW), and 0.1 μL of Taq DNA polymerase. For the foreground selection marker InDel7, PCR amplification was performed using a thermal cycler (Veriti, Applied Biosystems, Paisley, UK) with the following program: an initial denaturation at 94°C for 2 min; 35 cycles of denaturation at 95°C for 20 s, annealing at 60°C for 40 s, and extension at 72°C for 30 s; followed by a final extension at 72°C for 5 min and held at 4°C until analysis. (Kwon et al., 2012) For background selection, KASP (Kompetitive Allele-Specific PCR) amplification and allele discrimination were performed at the Seed Industry Promotion Center, Foundation of Agri. Tech. Commercialization and Transfer (Gimje, Korea), using the Nexar system (LGC Douglas Scientific, Alexandria, USA). KASP amplification was carried out according to the thermal cycling profile described (Cheon et al., 2018). After PCR amplification, fluorescence reading was performed for KASP genotyping.
Phenotypic evaluation and selection of advanced lines
In the BC2F2 generation, grain traits including thousand-grain weight (TGW), grain length (GL), grain width (GW), and the length-to-width ratio (LWR) were measured, and correlations among these traits were analyzed. Lines with high recurrent-parent genome (RPG) recovery and TGW ≥25 g were selected.
In the summer of 2020, seven lines were selected in the Miryang field based on RPG recovery, donor genome proportion, heterozygosity, residual chromosome segments, TGW, GL, GW, and LWR. During the 2021 summer season, agronomic traits such as heading date, culm length, panicle length, panicle number per plant, spikelet number per panicle, ripened grain ratio, TGW, brown rice percentage, and yield index were evaluated, and three promising lines were selected.
These three BC2F3 lines were further assessed for agronomic performance, disease resistance, and grain quality traits. Disease resistance evaluations included RSV, blast, and bacterial blight (BLB). Blast resistance was assessed under natural infection conditions at Miryang (Gyeongsangnam-do) and Sangju (Gyeongsangbuk-do), and BLB resistance was tested at Miryang using the leaf-clipping method with four Korean Xanthomonas oryzae pv. oryzae races (K1, K2, K3, and K3a).
Evaluation of RSV resistance
Forty rice seeds were sown in plastic trays (60 × 30 cm) and grown for two weeks. RSV-viruliferous small brown planthoppers (SBPH) at the second or third instar were then released into transparent plastic cages at a density of approximately seven insects per seedling for virus inoculation. Fourteen days after inoculation, the insects were removed, and the seedlings were transplanted to a glass greenhouse at the National Institute of Crop Science (NICS), Miryang, Korea. RSV resistance was evaluated one month after transplanting by calculating the percentage of healthy plants in each plot. Plants showing typical RSV symptoms, such as leaf curling, pale-green discoloration, and white stripe lesions, were classified as infected plants (Wang et al., 2021), whereas plants without these symptoms were considered healthy. The percentage of healthy plants was calculated as follows: Healthy plants (%) = (number of healthy plants / total number of plants) × 100 (Fang et al., 2013). Lines were classified as resistant when the proportion of healthy plants was ≥70%, whereas those with <70% healthy plants were classified as susceptible.
Statistical analysis
Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Tukey’s honestly significant difference (HSD) test, implemented in GraphPad Prism version 9.5.0 (GraphPad Software). Statistical significance was determined at p < 0.05. Groups sharing the same letter were not significantly different according to Tukey’s HSD test.
RESULTS
Development and segregation analysis of RSV-resistant lines
To introgress RSV resistance into Odae, segregation analyses were conducted from the BC1F1 to BC2F2 generations (Table 1). In the BC1F1 population, genotyping with InDel7 identified seven Odae-type homozygotes and three heterozygotes, which did not differ from the expected 1:1 ratio (χ2 = 1.60, p = 0.206). In the BC2F1 generation (n = 29), genotyping with InDel7 (Fig. S1) detected 11 homozygotes and 18 heterozygotes, again consistent with the expected 1:1 ratio (χ2 = 1.69, p = 0.194). In the BC2F2 population (n = 420), segregation also conformed to the expected 1:2:1 Mendelian ratio, with 87 resistant homozygotes, 224 heterozygotes, and 109 susceptible homozygotes (χ2 = 4.17, p = 0.124). After excluding 16 plants that died or showed poor early vigor, 71 resistant homozygous plants were transplanted in the 2019 summer season.
Table 1.
Foreground selection of backcross progenies using Indel7 markers linked to the RSV gene.
Recurrent parent genome recovery and background selection
The recovery rate of the recurrent RPG among the BC2F2 plants ranged widely from less than 70% to more than 90% (Fig. S2 and Fig. 2A). Background selection was performed using 116 polymorphic KASP markers that showed variation between Haedamssal and Odae, and heterozygous genotypes were detected at 56 marker loci. Based on these results, a second round of background selection was conducted for all 71 transplanted plants in the field. The average number of introgressed donor segments per line was 4.12, and the mean RPG recovery rate was estimated to be 79.8%. The mean proportions of donor-derived and heterozygous chromosomal segments were 9.0% and 11.2%, respectively, and the residual donor fragments were distributed across all 12 rice chromosomes.
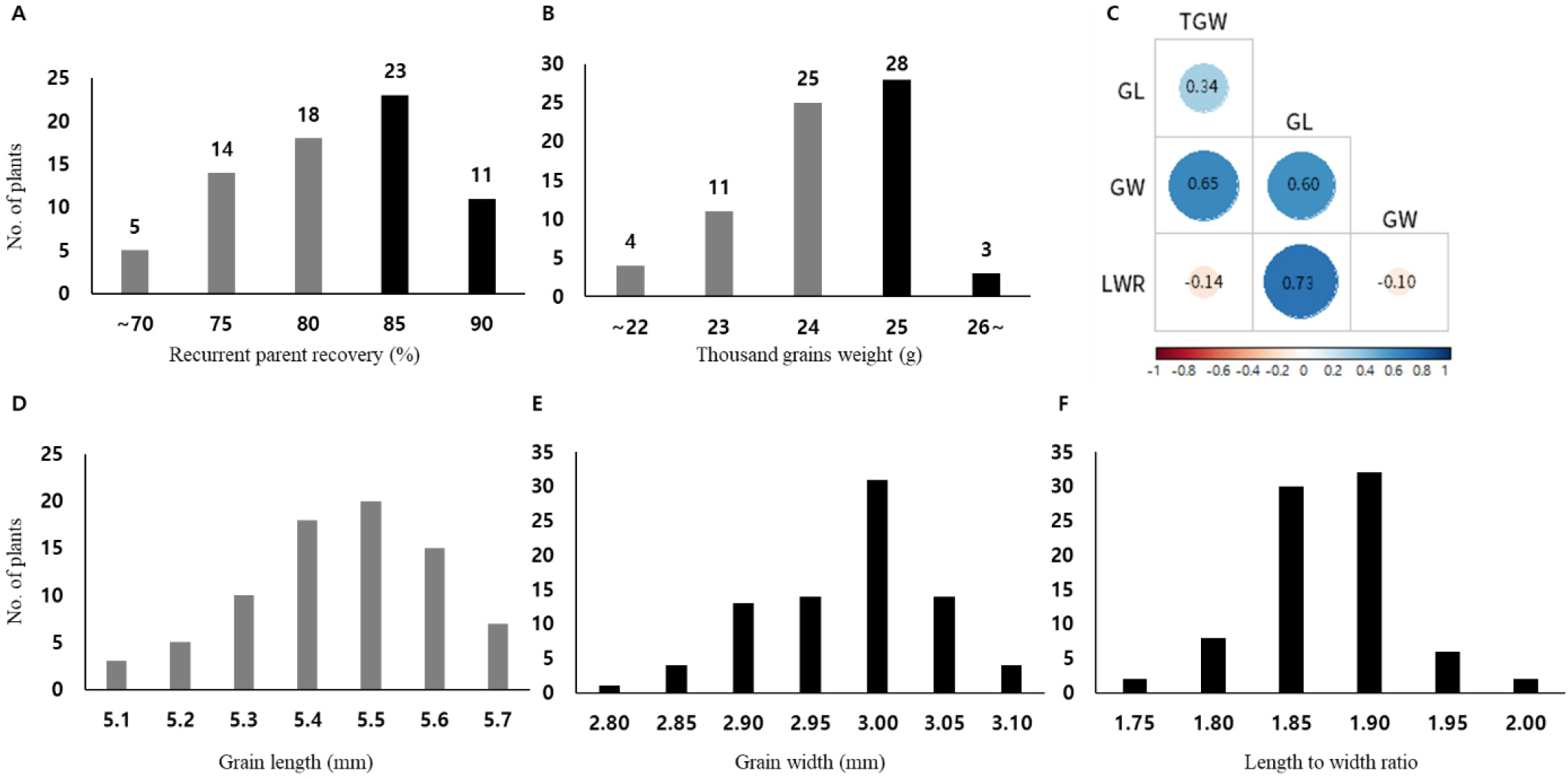
Fig. 2
Major phenotypic traits and correlation analysis of the NIL population. (A) Recovery rate of the recurrent genome in each NIL line, (B) Variation in thousand grain weight (TGW) among the NILs, (C) Correlation matrix showing the relationships among TGW, grain length (GL), grain width (GW), and length-to-width ratio (LWR), (D) Phenotypic variation in GL, (E) Phenotypic variation in GW, and (F) Phenotypic variation in the LWR.
Agronomic traits and correlation analysis
The TGW, one of the major yield components of Odae, exhibited a normal distribution ranging from 22 to 26 g (Fig. 2B). Thirty-one lines with an average TGW of ≥25 g, comparable to that of Odae, were selected. Correlation analysis (Fig. 2C-F) revealed a strong positive correlation between TGW and GW (r = 0.65) and a significant positive correlation between TGW and GL (r = 0.34), whereas a negative correlation was observed between TGW and LWR. A positive correlation was also detected between GL and GW, indicating a consistent covariation structure among grain size-related traits.
Development and characterization of the RSV-resistant NIL ‘Milyang397’
In the BC2F2 generation, seven lines were selected based on RPG recovery and grain traits and subsequently evaluated for agronomic performance (Table S1). Their heading date, culm length, and panicle length were comparable to those of the recurrent parent Odae. Further evaluation in 2021 reduced the population to three advanced lines, which were assessed for agronomic traits, disease resistance, and grain quality (Table 2 and 3). All three lines exhibited strong RSV resistance while retaining the major agronomic and grain quality characteristics of Odae. Among them, YR33079-1-3 showed an RPG recovery of 87%, exceeding the population mean of 79.8%. Donor-derived segments were primarily located on chromosomes 7 and 11, including the region harboring the Stvb-i resistance locus. Considering its favorable genomic composition and agronomic performance, together with slightly enhanced resistance to BLB, YR33079-1-3 was identified as the most promising line and subsequently designated as the RSV-resistant near-isogenic line ‘Milyang397’. Milyang397 displayed agronomic characteristics similar to those of Odae, including comparable grain size and shape (Fig. 3A-D), while exhibiting strong RSV resistance comparable to that of the donor parent Haedamssal in the seedling-stage bioassay (Fig. 3E).
Table 2.
Agronomic traits of Milyang397 and its selected sister lines.
Table 3.
Disease resistance and eating quality of Milyang397 and its sister lines.

DISCUSSION
RSV is a major viral disease that causes substantial yield losses in temperate East Asia, particularly in japonica rice. Early-maturing cultivars are often susceptible, which complicates efforts to combine early maturity with durable resistance. Consequently, developing NILs that support functional validation and facilitate the rapid introgression of RSV resistance genes into susceptible backgrounds is critical for sustainable rice production.
Traditional backcross breeding generally requires multiple backcrosses to minimize linkage drag and restore the recurrent parent genome, often taking more than four years to obtain the BC2F2 generation (Nandi et al., 2025). By contrast, the MABC approach can achieve high levels of RPG recovery within only two to three backcrosses. When combined with the RGA system, two to three generations can be advanced per year, reducing the total breeding time by more than two years (Das et al., 2022; Kang et al., 2019)
Marker-assisted background selection (MABS) has been reported as an effective strategy for minimizing linkage drag by rapidly restoring the recurrent parent genome and eliminating non-target donor segments (Miah et al., 2015). More than 90–95% genome recovery can typically be achieved at the BC2 or BC3 generation (Chukwu et al., 2020). The principle that lower donor fragment proportions improve the accuracy of estimating the net effect of target genes is also a key concept in NIL design (Hasan et al., 2015; Kim et al., 2021).
In this study, the RGA system was combined with the MABC strategy, and high-resolution background selection was conducted using 116 polymorphic KASP markers distinguishing Haedamssal and Odae. Heterozygous genotypes were detected at 56 marker loci, with an average of 4.12 donor fragments per line. The mean RPG recovery rate was 79.8%, indicating that high-recovery lines could be obtained within only two years at the BC2F2 stage. Although the mean RPG recovery rate at the BC2F2 stage was 79.8%, the distribution was wide, and a subset of lines exceeded 90% RPG recovery. This variation enabled the selection of high-recovery NILs with minimal donor background while retaining the target resistance allele (Semagn et al., 2006; Stam & Zeven, 1981). These results showed a similar trend to previous studies on rice MABC and RGA-based generation acceleration (Kang et al., 2019; Kim et al., 2021), suggesting that combining RGA and MABC effectively accelerates recurrent genome recovery and facilitates efficient introgression of resistance genes.
The newly developed line ‘Milyang397’ exhibited stable expression of RSV resistance while maintaining key agronomic traits such as heading date, culm length, and panicle length comparable to those of the recurrent parent Odae. This demonstrates that the NIL/MABC approach enables the functional validation of target genes while preserving the inherent agronomic characteristics of the recurrent parent (Kamal et al., 2023; Khanna et al., 2015). In addition, molecular marker–based selection is not affected by inoculation intensity or environmental conditions, making it advantageous for accurate and reproducible evaluation of RSV resistance (Collard & Mackill, 2008).
The high-recovery NILs developed in this study exhibited stable expression of RSV resistance. Furthermore, introgression of the resistance gene did not negatively affect growth or yield-related traits. This suggests that the RSV resistance gene is not linked to growth-suppressing loci, and its introgression does not compromise the productivity of early-maturing cultivars. Furthermore, the consistency between MAS results using the InDel7 marker and actual RSV resistance expression demonstrates that this marker is highly reliable for the precise tracking and selection of the RSV resistance gene. In contrast to previous studies that primarily introgressed RSV resistance into medium- or late-maturing genetic backgrounds, this study demonstrates the efficient development of an RSV-resistant NIL in an early-maturing japonica background through the combined use of MABC and RGA. These results suggest that this strategy can facilitate the rapid development of RSV-resistant cultivars adapted to early-maturing japonica rice breeding programs in Korea. Overall, these results confirm that an efficient NIL platform was successfully established, enabling unbiased evaluation of RSV resistance effects in an early-maturing background and providing valuable genetic resources for elucidating RSV resistance mechanisms and breeding resistant rice cultivars in the future.
CONCLUSION
This study demonstrates that the combination of marker-assisted backcrossing (MABC) and a rapid generation advance (RGA) system is an effective approach for developing rice near-isogenic lines with resistance to rice stripe virus (RSV). Using Haedamssal as the donor parent and Odae as the recurrent parent, RSV resistance conferred by the Stvb-i gene was successfully introgressed while maintaining the major agronomic and grain quality traits of the recurrent parent. Background selection using polymorphic KASP markers enabled efficient recovery of the recurrent parent genome at the BC2F2 generation, thereby reducing the time required for line development. The selected NIL, ‘Milyang397’, exhibited stable RSV resistance comparable to that of the donor parent without adverse effects on growth or yield-related traits. These results indicate that the RGA–MABC strategy provides a reliable framework for the rapid development and evaluation of RSV-resistant rice lines and can support the breeding of resistant cultivars in early-maturing japonica backgrounds.
