

서 론
야생 호밀(wild rye)인 Leymus racemosus (2n = 4x = 28, NsNsXmXm)는 해변이나 건조한 지역, 그늘진 습한 숲에서 많이 자생하며 다양한 유전적 유용변이를 가지고 있다(Fan et al., 2009). Leymus종은 염해(鹽害)와 가뭄에 강하며 꽃이 피는 5월과 6월 개화기(開花期)에 발생할 수 있는 고온 현상에 대한 저항성을 가진 유전인자가 있으며, 생물학적 질화 작용을 억제(biological nitrification inhibition, BNI)하며 백색 녹병, 붉은 곰팡이병에 대한 저항성을 가지고 있다(McGuire&Dvorak, 1981; Chen et al., 2005; Subbarao et al., 2007; Mohammed et al., 2014). 야생 호밀의 작물학적 유용인자들은 밀 품질 향상을 위하여 야생 호밀 염색체와 밀 염색체간 전좌(translocation)가 일어난 계통을 육성하였으며, 이 밀 전좌 계통은 붉은 곰팡이병에 저항성이 가지고 있는 것을 연구를 통하여 확인하였고, 야생 호밀 염색체를 핵산표지인자를 이용하여 각 염색체를 확인하는 연구가 최근까지 수행되었다(Chen et al., 2005; Edet et al., 2018). 야생 호밀의 각각의 염색체 특성을 이용하기 위하여, 6배체 보통 밀(Triticum aestivum L. cv. Chinese spring, 2n = 6x = 42, AABBDD)의 유전적 배경(genetic background)에 야생 호밀의 한 종인 Leymus racemosus (2n = 4x = 28, NsNsXmXm)로부터 각 한 쌍의 상동염색체(homologous chromosome)가 첨가된 외래 이가 염색체 첨가 계통(disomic addition lines, DAs)이 육성되었다(Kishii et al., 2004).
야생 호밀 염색체 첨가 계통의 특징은 이삭의 형태이다. 야생 호밀 염색체 첨가계통의 이삭 형태는 각 한 쌍의 상동염색체에 의해 표현형의 차이를 보인다(Kishii et al., 2004). 야생 호밀은 보통 밀보다 긴 이삭을 가진 것이 특징이지만, 각 염색체 첨가 계통의 이삭의 길이는 가장 짧은 이삭 길이가 보통 밀의 절반정도 수준이며, 그 외 염색체 첨가 계통의 이삭길이는 유전적 배경이 보통 밀이기 때문에 전반적으로 보통 밀과 유사한 수준을 보인다. 하지만 영화의 수와 형태가 첨가된 야생 호밀 염색체에 따라 고유한 특성을 가진다(Kishii et al., 2004). 또한, 종실의 폭과 길이 및 모양에도 야생 호밀 염색체가 영향을 주는 것으로 보이나 이런 특성과 야생 호밀 염색체사이의 연관관계에 대한 연구는 매우 미흡한 실정이다.
본 연구에서 야생 호밀의 염색체가 첨가된 밀 계통의 단백질 발현 양상과 이차원전기영동(2-DE)을 이용하여 단백질 발현을 비교 분석하였고, 질량 분석을 통하여 단백질의 기능을 확인하여 해석한 결과를 보고하고자 한다. 이를 통하여 야생 호밀의 유용 형질이 보통밀의 생물학적 및 비생물학적 스트레스에 저항성에 대한 작물학적 유용 가치를 향상시킬 수 있는데 기여하고자 한다.
재료 및 방법
공시재료의 염색체 관찰
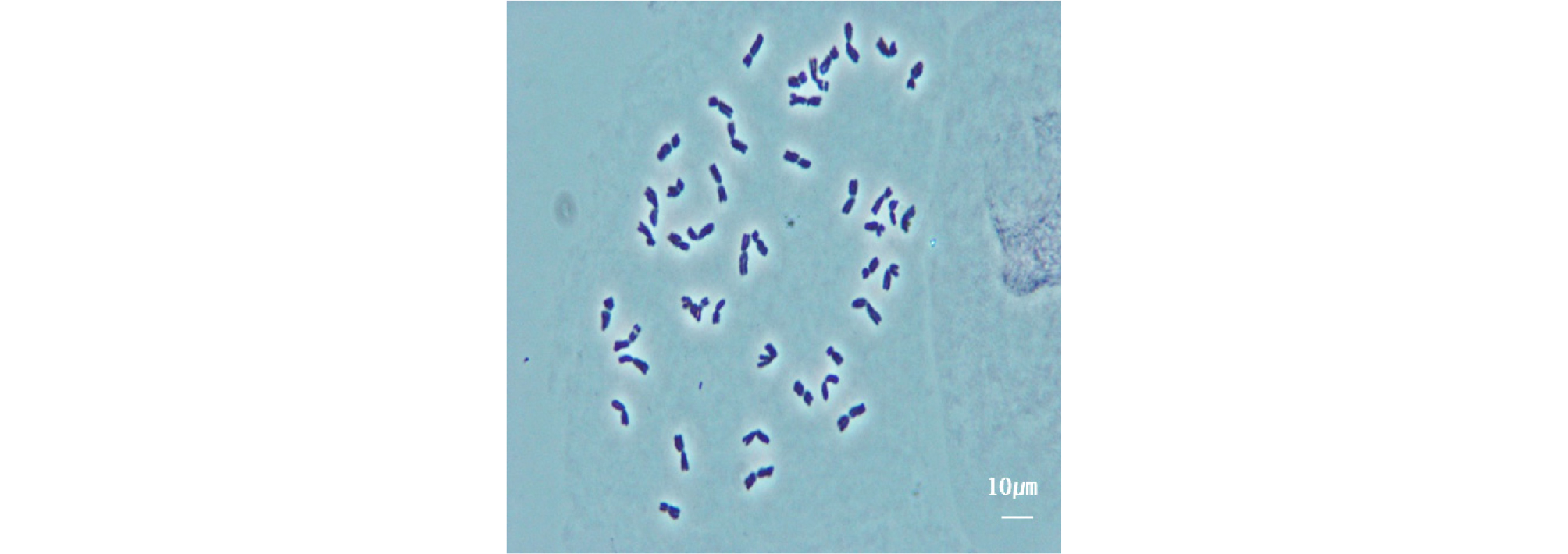
보통밀인 Chinse spring (CS)에 Leymus racemosus 가 첨가된 9개 DAs은 일본 돗토리대학 히사쉬 쯔지모토 교수가 감사하게도 제공해 주었다. 이들 DAs의 염색체 관찰을 위하여 뿌리(root tip)의 분열세포(meristem cell)에서 체세포(mitotic cell)를 2% acetocamine 용액으로 염색한 후에 염색체를 squash 방법(Östergren&Heneen, 1962)으로 체세포 슬라이드를 만들어 위상차 현미경(Olympus Bx53F2, Japan)으로 관찰하였다. DAs 9계통의 염색체 수는 42개의 밀 염색체와 2개의 외래 염색체로 총 44개의 염색체로 구성되어 있는 것을 확인하였다(Fig. 1).
단백질 추출 및 정량 분석
액체질소로 곱게 마쇄한 시료 0.1g에 Urea (4.8 g), Np-40 (0.2 mL), ampholine (0.2 mL), 2-Mercap (0.5 mL), 및 PVP- 40 (0.5 g)로 구성된 단백질 추출 용액(lysis buffer) 1 mL를 첨가하여 반응시킨 후, 4°C, 15,000 rpm에서 15분간 원심분리하였다. 원심분리 후, 상등액을 단백질 정량분석에 이용하였다. 단백질 농도 측정은 Lowry et al. (1951)의 방법에 준하여 RC DC protein assay를 사용하여 진행하였으며, Reagent A1은 DC Reagent A와 DC Reagent S를 50 : 1로 희석하였고, 단백질 농도 측정은 750 nm에서 분광광도계(SHIMADZU, UV-1700)를 이용하였다.
단백질 발현 분석
CS와 9개 DAs의 단백질 발현 분석은 SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis)와 2-DE (2-dimensional electrophoresis)를 이용하였다. SDS-PAGE는 Laemmli (1970)의 방법에 준하여 수행하였는데, SDS 겔은 12% separating gel (1.6 mL DDW, 2.0 mL 30.0% acrylamide solution, 1.3 mL 1.5M Tris-HCl (pH 8.8), 50 µL 10% SDS, 50 µL 10% Ammonium persulfate, 2 µL TEMED)과 5% stacking gel (1.4 mL DDW, 330 µL 30.0% acrylamide solution, 250 µL 1.0M Tris-HCl (pH 6.8), 20 µL 10% SDS, 20 µL 10% Ammonium persulfate, 2 µL TEMED)을 이용하였며, 전기영동 후, CBB (coomassie brilliant blue) G-250로 염색하였다. 2-DE는 등전점(isoelectric point, pI)을 이용한 1차 전기영동 겔은 Urea (4.8 g), 30% Acrylamide solution (1.6 mL), 10% NP-40 (2 mL), Ampholyte pH 3-10 (0.25 mL), 10% APS (15 µL), 및 TEMED (10 µL)로 만들었으며, 2차 전기영동의 겔 조성은 SDS-PAGE와 동일하였다.
단백질 기능 확인
CS와 9개 DAs의 특이 단백질 발현을 확인하기 위하여 MALDI-TOF/MS (AXIMA CFR+ Plus, Shimadzu, Japan)을 이용하였다. 단백질 동정은 질량분석기로부터 분석한 샘플의 MS spectra data 값을 먼저 MASCOT (Matrixscience, London UK http://www.matrixscience.com)의 database에서 National Center for Biotechnology Information (NCBI, https:// www.ncbi.nlm.nih.gov)를 이용하였다. 실험과정에서 변성이 일어난 단백질의 동정 보완을 위해 search parameters를 acetyl (K), carbamidomethyl (C), oxidation (M), propionamide (C) 선택하였고, peptide의 오차범위는 100 ppm 안에서 동정하였다. NCBI 및 Universal Protein Resource (UniPROT, https:// www.uniprot.org)의 database를 통한 protein name과 accession number의 정보를 Protein Information Resource (PIR, https:// proteininformationresource.org)에서 cellular component, molecular function 및 biological process 같은 단백질 정보를 수집하여 protein description과 peptide sequence의 정보로 protein sequence를 찾아 동정된 단백질의 기능별 분류에 이용하였다(Wu et al., 2003).
결과 및 고찰
단백질 발현 양상 비교
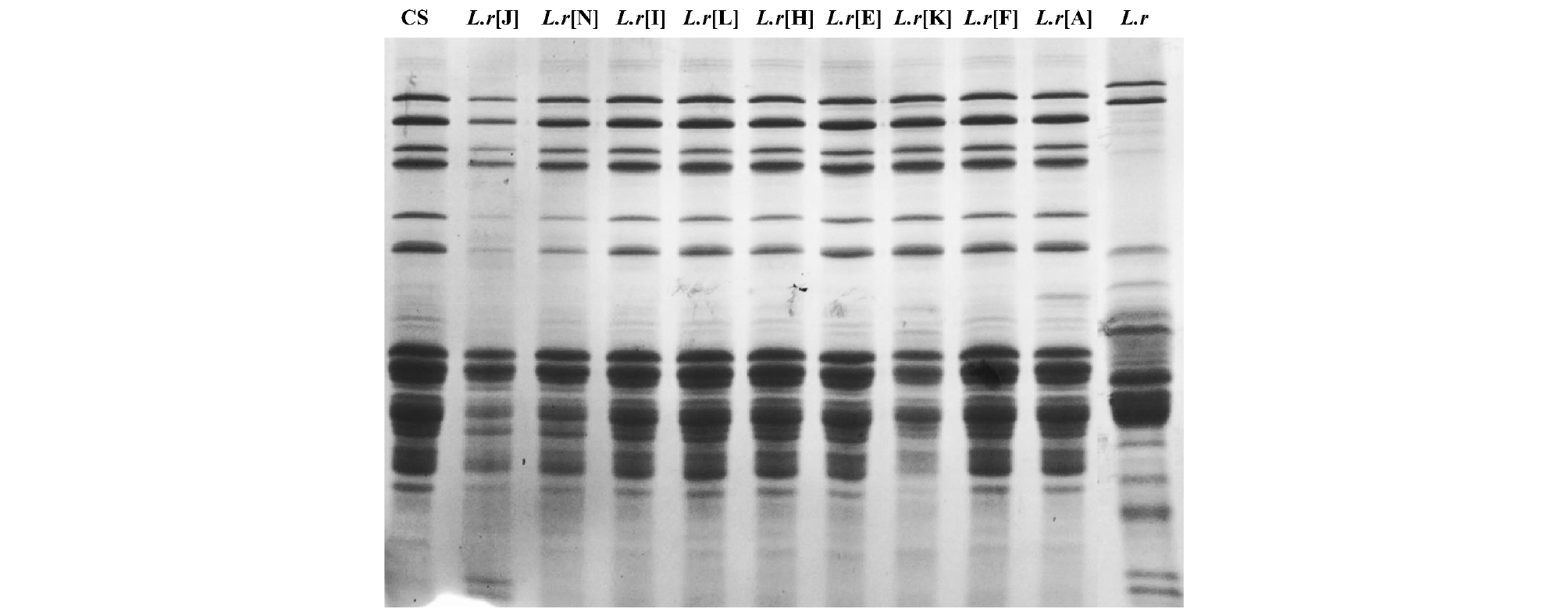
CS와 9개 DAs의 SDS-PAGE의 HMW-GS 발현 양상 비교 결과(Fig. 2), CS의 고분자 글루텐 서브유닛(high molecular weight glutenin subunit, HMW-GS)은 알려진 것처럼 Glu-Alc, Glu-Blb와 Glu-D1a로 4개의 단백질이 발현되었으며, 9개 DAs의 HMW-GS도 Chinese spring과 같은 단백질 밴드 패턴을 보였지만, 야생 호밀과는 달랐다. 저분자 글루텐 서브유닛(low molecular weight glutenin subunit, LMW- GS)의 발현 양상도 Chinese spring과 9개 DAs은 유사했지만 야생 호밀은 다른 패턴을 보였다. LMW-GS는 HMW-GS보다 복잡한 발현 양상을 보이기 때문에 SDS-PAGE를 이용한 분석에는 한계가 있지만(Anderson et al., 2009), DAs중에서 L.r[J] 계통은 야생 호밀에서 발현되는 15~20 kDa의 2개 단백질이 나타난 것을 확인하였다. 또한, SDS-PAGE 상에서 HMW-GS과 LMW-GS 사이에 발현되는 것으로 보이는 ω-gliadine은 CS와 9개 DAs에서 발현되었고, 야생 호밀에서는 나타나지 않았다.
이차원전기영동(2-DE)을 이용한 단백질 발현 결과(Fig. 3), 9개 DAs은 CS와 유사한 발현 양상을 보였지만, 단백질 발현 정도(degree of protein expression)는 차이를 보였다(Table 1). 이미지 분석으로 CS의 단백질 발현 정도를 기준으로 동일한 위치의 단백질 스팟의 발현 정도를 비교한 결과, L.r[A]는 단백질 발현이 증가(up regulation)한 스팟은 51개였으며, 44개 스팟은 감소(down regulation)하였고, L.r[E], L.r[F], L.r[H], L.r[I], L.r[J]와 L.r[K]의 단백질 발현 증가 스팟은 각각 54개, 58개, 52개, 46개, 55개, 51개와 51개였고, 단백질 발현 감소 스팟은 각각 37개, 43개, 49개, 40개, 44개, 44개와 47개였다. 단백질 발현 정도가 증가된 단백질 스팟이 가장 많은 계통은 L.r[F] 계통이었으며, 가장 적은 계통은 L.r[N]였다. L.r[I]은 다른 DAs과 다르게 단백질 발현 정도가 CS보다 단백질 발현이 감소되는 스팟이 증가되는 스팟보다 많았다. 이미지 분석을 통한 단백질 발현 양상의 비교는 단지 야생 호밀 한 쌍의 염색체가 첨가됨에 따라 단백질의 발현 정도에 차이를 비교하였을 뿐 첨가된 야생 호밀의 염색체가 어떤 과정을 통해 단백질의 발현 정도에 영향을 미치는지는 좀 더 많은 분석과 연구가 필요하다.
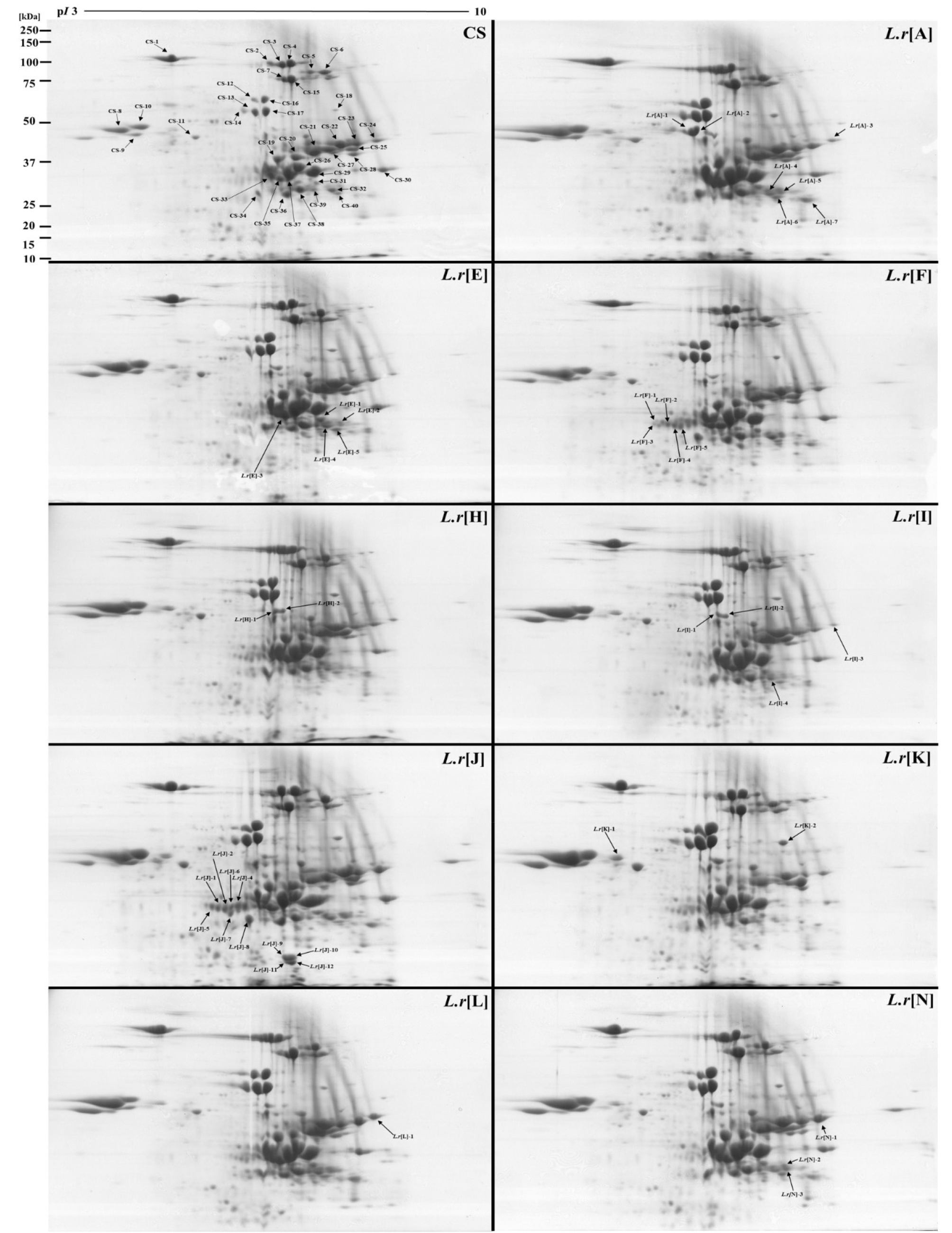
Table 1. Comparison of the protein expression degrees according to normal volume by image analysis.*
단백질 기능 해석
질량 분석을 이용하여 CS는 38개 L.r[A]은 5개, L.r[E]은 5개, L.r[F]은 4개, L.r[H]은 2개, L.r[I]은 4개, L.r[J]은 7개, L.r[K]은 1개, L.r[N]은 3개로 총 67개 단백질이 동정되었다(Table 2). Protein Information Resource (PIR, https://pro teininformationresource.org)에서 단백질의 기능을 확인하여 mascot score가 가장 높은 것만 선택하여 총 11개의 그룹으로 분류되었다(Wu et al., 2003).
Table 2. Identification of protein function by MALDI-TOF MS.
2)MW, calculated molecular weight
3)pI, calculated isoelectric point
첫번째 그룹은 스트레스 반응에 관련된 단백질로 CS에서 4개의 스팟(CS-1, -9, -10, -30)이 여기에 속했으며, L.r[J]에서 한 개의 스팟(L.r[J]-10)이 해당되었다. CS-1와 CS-9는 L-2와 관련 있고, L.r[J]-10는 Mol1 단백질의 부분 염기서열이 일치하였고, CS-10과 CS-30는 아직 기능이 밝혀지지 않은 단백질로 나타났다. L-2 단백질(late protein L2)은 항균성 단백질의 전구체(precursor of antimicrobial peptides)로 알려져 있으며, 곰팡이 균에 대한 방어 기작에 관련된 단백질로써 밀의 발아시 중요한 역할을 하는 단백질로 알려져 있다(Utkina et al., 2012). Mol1 단백질은 protein MLO family의 영역에 있는 단백질로 밀에서 동정되었으나 연구가 미흡하며, 단지 병원체에 대한 저항성과 잎 세포사(cell death)와 관련된 것으로 추정하고 있으며, 밀의 흰가루병(powdery mildew fungus)에 대한 광범위 저항성과 관련된 것으로 보고 있다(database of UniProt, https://www.uniprot.org/uniprot/ P93766). 밀의 흰가루병은 확산이 빠르며, 발생 정도가 심하기 때문에 흰가루병 저항성 향상에 유용할 것으로 생각한다. 하지만 밀의 병저항성 향상을 위해서는 Mol1 단백질에 대한 집중적인 연구가 필요하다.
두번째 그룹은 DNA 및 RNA 생성에 관여하는 전사인자(transcription factor, TF) 단백질로써 유전자 발현에 있어 매우 중요한 역할을 하고 있다(Latchman, 1993). 전사인자와 관련된 단백질 스팟은 CS-2, CS-25, Lr[H]-1과 Lr[N]-1인데, WRKY transcription factor (CS-2), heat shock factor B1b (CS-25), isopentenyl transferase (ipt) (Lr[H]-1), low- molecular-weight glutenin subunit (Lr[N]-1)]로 확인되었다. WRKY transcription factor는 오직 식물에 발견되는 전사제어인자의 가장 큰 그룹 중 하나이다(Rushton et al., 2010). 야생 호밀 염색체 첨가 계통에서 up-regulation을 보인 Lr[H]-1, isopentenyl transferase (ipt)는 식물의 병원성균에서 추출된 유전자로 cytokinin 생합성하는 과정에서 촉매 작용을 돕고, 스트레스 저항성을 향상시키며, 다년생의 잡초를 포함한 다양한 식물에서 고온 저항성을 가진다(Guo et al., 2010; Xu et al., 2009). CS-25, HSP (heat shock protein)단백질군은 고온스트레스와 정의 상관관계가 있어 HSP들은 고온스트레스로부터 세포를 보호하며 HSP의 축적은 고온저항성을 향상시키는 것으로 알려져 있으며 대개 HSPs (small heat shock proteins), HSP 60, HSP 70, HSP 90, HSP 100등 다섯가지로 분류한다(Lamb et al., 2003; Wang et al., 2004; Dzaman-Serafin et al., 2005). 또한, HsfB1b 유전자는 DNA-binding transcription factor activity와 sequence- specific DNA binding의 역할을 한다고 알려져 있다(Kadonaga&Tjian, 1986). 야생 호밀은 주로 중앙아시아(central Asia)와 동유럽(eastern Europe)의 해안가 주변에 분포하고 있다(Kishii et al., 2004). 이러한 열악한 환경에서의 적응성(adaptation)은 기후변화에 대응하여 밀 품질 향상에 도움이 될 것으로 생각한다. 우리나라는 서해안의 신두리 해안에서 갯그령이라 불리는 야생 호밀종(Leymus mollis)이 분포하고 있다. 이러한 야생 호밀은 열(heat)에 대한 저항성 뿐만 아니라 염(salt)과 건조(drough)에 대한 저항성이 있는 것으로 알려져 있다(McGuire&Dvorak, 1981). 따라서, 국내 야생 호밀인 갯그령을 이용한 연구가 필요할 것으로 생각한다.
Lr[N]-1, LHW-GS (low molecular weight glutenin subunit) 단백질은 저분자 글루테닌 서브유닛이로 밀 품질에 관련이 있다(Masci et al., 1998). 글루텐(Glutein)은 밀의 품질을 결정하는 중요한 단백질이다(Apples&Lagudah, 1990). 밀은 다른 작물과는 다르게 단백질의 함량에 의해 밀가루의 종류가 구별되며, 이러한 단백질은 최종 가공품의 가공적성 뿐만 아니라 품질에도 영향을 미친다(Dhaka&Khatkar, 2015). 이 단백질은 야생 호밀의 LMW-GS으로 밀의 가공적성 향상에 어떻게 영향을 미칠지 고성능 액체 크로마토그래프(high- performance liquid chromatography, HPLC) 등을 이용하여 밀의 LMW-GS와 단백질 분자량 뿐만 아니라 특성 비교 및 실질적인 품질 평가가 필요할 것으로 생각한다.
당분해(glycolysis) 관련 단백질은 CS-29, 12-oxo-phytodienoic acid reductase (CS-29)과 CS-37, glyceraldehyde- 3-phosphate dehydrogenase (GADPH)로 확인되었다. CS-29, 12-oxo-phytodienoic acid reductases (OPRs)는 OPRI과 OPRII로 분류되며, 자스몬산(jasmonic acid) 생합성 과정에 관여한다(Dong et al., 2013). OPR(s)는 old yellow enzyme (OYE) 단백과 유사한 서열을 가지고 있으며, 항산화 및 산화스트레스와 관련이 있다고 연구를 통해 밝혀지고 있다(Fitzpatrick et al., 2003; Dong et al., 2013). CS-37, GADPH는 광합성 중 탄소 동화 작용(carbon dioxide assimilation)과 광합성 암반응 Calvin-cycle에 작용하는 효소로 기본적인 세포 이화작용 과정(catabolic processes)을 제공하는 단백질이며, 에너지 생산 및 광합성(photosynthesis)에 매우 중요한 역할을 하는 효소로 당분해에 연관이 있다(Price et al., 1995; Sirover, 1997; Marri et al., 2005).
세포 기관 구성(organelle organization) 관련 단백질은 Lr [E]-1, actin-depolymerizing factor (ADF) 7과 Lr[J]-4, rps2로 확인되었다. Actin depolymerizing factor의 기능은 미세섬유(microfilament) 단백질에 속하며 액틴 미세섬유(actin microfilament)의 중합체를 단위체로 하는 해중합화(depolymerization)를 돕는 역할을 한다. 최근 밀에서 ADF와 녹병 병원균에 대한 저항성과의 관계에 대한 연구가 수행되었다(Zhang et al., 2017).
단백질 수송(protein transport) 관련 단백질은 CS-36, truncated putative heavy metal transporter variant 2, Lr[A]-5, truncated putative heavy metal transporter variant 1과 Lr[E]-4, Ran-binding protein로 확인되었다. 수송에 관련된 단백질 중 Ran-binding protein (Lr[E]-4)은 GTP (guanosine triphosphate)-binding nuclear protein으로 널리 알려져 있으며, 염색질 응축(chromatin condensation)과 세포 주기와 GTP-결합 단백질의 세포내 수송에 관여한다(Moore, 1994).
번역(translation) 관련 단백질은 Lr[A]-6, Chain I, Cryo- Em Structure Of The Mammalian Sec61 Complex Bound To The Actively Translating Wheat Germ 80s Ribosome로 확인되었다. 복제(reproduction) 관련 단백질은 Lr[F]-3, beta- expansin TaEXPB1로 확인되었다. Expansin은 식물의 세포벽에 발견된 비효소적 단백질이며, beta-expansin TaEXPB1은 밀의 소포자(microspores)에서 분리 동정되었으며, 유성생식에 관여하는 것으로 알려져 있다(McQueen-Mason et al., 1992; Jin et al., 2006, https://www.uniprot.org/uniprot/Q56TQ1). 광합성(photosynthesis) 관련 단백질은 Lr[A]-7, putative oxygen-evolving complex precursor와 CS-17&CS- 18, ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit로 확인되었다. 광합성은 식물이 대사과정에서 필수적인 역할을 하며, 환경적인 스트레스에 영향을 받기 쉽다. 본 연구를 통해 확인한 oxygen-evolving complex precursor (Lr[A]-7) 단백질은 카드뮴(cadmium, Cd) 스트레스를 주었을 때 down-regulation 되는 것이 확인되었다(Kieffer et al., 2008).
산화 환원 항상성(redox homeostasis) 관련 단백질은 Lr[N]- 2, thioredoxin H로 확인되었다. Thioredoxins은 작고 어디에나 존재하는 단백질로써, 이황화결합(disulfide bond)을 환원시킬 수 있는 생리학적 과정에 중요한 역할을 한다(Prinz et al., 1997). 밀에서는 저장 단백질인 글리아딘(gliadins)과 글루테닌(glutenins)이 thioredoxin H에 의하여 감소되는 연구결과가 Kobrehel et al. (1991)에 의하여 보고되었다. 단백질 증첩과 안정(protein folding and stabilization) 관련 단백질은 CS-33, calreticulin (CRT)로 확인되었고, 소포체에 존재하는 칼슘결합단백질(Ca2+-binding protein)로 식물에서는 내건성(drought resistance)과 관련된 것을 증명하기 위하여 밀에서 TaCRT 단백질이 동정되었으며, 밀의 TaCRT는 다른 식물의 CRT(s)와 매우 높은 상동성을 보였고, TaCRT 전사는 건조 스트레스에 대응하여 파종시기에 축적되는 것으로 확인되었다(Jia et al., 2008).
물질대사(metabolism) 관련 단백질은 glycosyltransferase, HGA-like, putative, expressed (CS-7&CS-13), calcium- dependent protein kinase (CS-16), ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (CS-14, CS-24, Lr[A]-1, Lr[A]-2), ubiquitin-protein ligase, putative, expressed (Lr[H]-2), serine/threonine protein kinase (CS-11), heterotrimeric G protein alpha subunit (CS-22), SHAGGY-like kinase (CS-21), trehalose phosphate phosphatase, partial (Lr[F]-1), (1,4)-beta-xylan endohydrolase (Lr[I]-2), gamma tocopherol methyltransferase (Lr[J]-5), phytoene synthase 1 (Lr[K]-1), S-phase kinase protein-like protein (Lr[N]-3)로 다수의 인자가 물질대사와 관련된 단백질로 확인되었다.
현재까지 야생 호밀에 대한 작물학적 이용 가치에 대한 연구는 매우 미흡하며 이종교배를 통하여 얻은 유용유전자원에 대한 연구 역시 매우 미흡한 실정이다. 향후 고성능 액체 크로마토그래피(high-performance liquid chromatography, HPLC)와 같은 고처리 탐색(high-throughput screening)을 통해 좀 더 면밀한 분석이 필요하며, 국내에서도 중국과 일본처럼 이종교배를 통한 외래 염색체 첨가 밀 계통을 환경변화 대응 및 지속가능한 농업을 위한 작물학적 가치 증진에 대한 적극적인 연구가 필요하다.
적 요
야생 호밀 염색체 첨가 계통의 단백질 발현 양상을 보통 밀과 비교함으로써 발현의 차이를 보이는 단백질의 기능을 동정함으로써 야생 호밀의 작물학적 유용 가치를 확인하고자 이 연구를 수행하였다. 전반적으로 야생 호밀 염색체 첨가계통은 보통 밀의 유전적 배경을 바탕으로 건조와 열에 대한 비생물학적 스트레스에 대한 저항성 관련 단백질과 바이러스성 병원균에 대한 저항성 관련 단백질 및 척박한 환경에 적응하는 생리대사에 관련된 단백질을 가지고 있는 것을 확인하였다. 하지만 아직 야생 호밀의 단백질 기능에 대한 정보와 작물학적 이용에 대한 연구가 미흡한 상태이다. 앞으로 국내 야생 호밀의 유용 유전자원으로써의 작물학적 이용과 기능에 대한 지속적인 연구가 필요하다.
