

최근의 지구온난화로 인한 이상기후가 자주 발생함에 따라 작물환경에 영향을 미치고, 특히 고온에 의한 작물의 심각 한 피해가 예상된다(Sadat et al., 2013). 고온스트레스는 저 온, 염스트레스 등과 같이 주요한 환경스트레스 중 하나로 서 작물의 생육에 영향을 끼쳐 최종적으로 곡물의 수량을 감소시킨다(Boyer, 1982). 특히 밀의 생식단계 또는 개화 후 성숙되는 기간의 고온스트레스는 심각한 수량 감소를 일으 킬 수 있다(Hays et al., 2007). Gibson and Paulsen (1999) 등은 개화 후 10일에서 성숙 단계 동안에 35℃의 고온 피해 를 받으면, 수량 78%, 종자 수 63%, 종자무게가 29% 정도 감소된다고 보고하였다. 그러나 개화 후 15일에서 완전 성 숙 때까지의 고온은 종자무게만 18% 감소시킨다고 보고되 었다(Gibson and Paulesn, 1999). 고온의 영향으로 최적온도 에서 평균온도가 1℃ 상승하면, 세계적으로 곡물생산의 3~4% 의 감소가 발생한다고 보고되었다(Wardlaw et al., 1989). 비록 고온스트레스에서도 생산안정성을 보이는 여러 품종 들이 확인되고 있지만, 품종 육성 단계에서 육종가가 현장 에서 내고온성 등과 같은 환경스트레스와 관련된 양적 형질 을 효과적으로 선발하는 데는 어려움이 따른다(Hays et al., 2007; Yang et al., 2002).
단일 목적 형질을 위한 몇몇 양적형질 유전자좌(Quantitative trait loci, QTL)가 존재하기 때문에 목적형질과 밀접하게 연관되어 있는 분자마커 또는 실제 유전자 염기서열을 이용 한 간접적 선발의 효율이 표현형적 선발보다 시간적, 경제 적으로 효율이 높을 수 있다. 이론에 기반을 둔 시뮬레이션 연구를 통해 다원발생 형질(polygenic trait) 선발을 위해서 는 전통적으로 수행되는 표현형적 선발보다 분자표지이용 선발(Marker assisted selection, MAS)의 효과가 더 클 수 있다는 것이 제안되었다(Lande and Thompson, 1990; Zhang and Smith, 1992). 분자표지이용 선발은 주로 큰 집단에서 목적형질의 유전율이 낮을 때 이용할 수 있다. 또한 분자마 커가 목적하는 양적형질(QTL)과 밀접하게 연관(< 5.0 CM) 되어 있을 때 효율이 높고, 특히 선발은 분자마커와 양적형 질 사이에서 재조합이 일어나기 전인 초기 세대에서 수행되 는 것이 좋다. 많은 연구를 통해 목적형질의 염색체상 유전 자 위치 등과 다양한 DNA 마커가 보고되어 있으며, 현재 CIMMYT 등 국외 여러 기관에서 밀의 고단백질(Distelfeld et al., 2006), 밀가루색(Sharp et al., 2001), 종자의 경도(Giroux and Morris, 1997)와 반죽 강도 및 볼륨(Mclauchlan et al., 2001) 등 품질 관련 DNA 마커와 붉은곰팡이병(Fusarium head blight) (Anderson et al., 2001)과 녹병(Stem rust) (Prins et al., 2001) 등의 연구에 MAS를 적용하고 있다. 밀의 유 용형질에 대한 유전적 특성을 이용한 다양한 분자마커 시스템 이 사용되고 있는데 그 중에서 microsatellite (Simple sequence repeat, SSR)는 공우성 마커로서 유전분리 초기 세대에서 유용하게 사용할 수 있다(Chen et al., 1998; Temnykh et al., 2001). SSR은 Restriction fragment length polymorphism (RFLP) 이나 Random amplification polymorphic DNA (RAPD)에 비해 재현성과 확실한 다형성을 확인할 수 있으며, 유전자 마커 개발의 기본적인 자원으로 매우 중요하다(Sadat et al., 2013). 이러한 SSR은 밀의 종간 유연관계 분석 등에 사용 이 되었으며, 현재 형질과 관련된 유전자 지도 작성 및 마커 개발에 많이 사용되고 있다(Houghton et al., 2001). 따라서 본 연구는 기존의 밀의 고온스트레스와 관련된 다양한 SSR 마커 중에서 최근 연구를 통해 가장 연관이 있는 것으로 보 고된 마커(Table 2)를 이용하여 국내에서 육성된 품종과 터 키에서 육성 중인 내고온성 유망 계통을 평가하였다.
Table 2.
Characteristics and genotypes of SSR markers for marker assisted selection of 31 wheat varieties for heat stress resistance.
| Marker name | Forward sequences (5‘→3’) | Reverse sequences (5‘→3’) | Chromosomal location | QTLs | Number of allels | PIC value | Reported by |
|---|---|---|---|---|---|---|---|
| Gwm11 | GTGAATTGTGTCTTGTATGCTTCC | GGATAGTCAGACAATTCTTGTG | 1B | Grain-filling duration | 4 | 0.62 | Yang et al. (2002) |
| Gwm291 | AATGGTATCTATTCCGACCCG | CATCCCTAGGCCACTCTGC | 5A | HIS Single kernel | 16 | 0.83 | Mason et al. (2010) |
| Gwm268 | TTATGTGATTGCGTACGTACCC | AGGGGATATGTTGTCACTCCA | 1B | HIS Single kernel | 5 | 0.71 | Mason et al. (2010) |
| Gwm293 | TCGCCATCACTCGTTCAAG | TACTGGTTCACATTGGTGCG | 5A | Grain-filling duration | 10 | 0.83 | Yang et al. (2002) |
| Gwm325 | TTTTTACGCGTCAACGACG | TTTCTTCTGTCGTTCTCTTCCC | 6D | Grain-filling duration | 4 | 0.69 | Mason et al. (2010) |
| Gwm484 | AGTTCCGGTCATGGCTAGG | ACATCGCTCTTCACAAACCC | 2D | Grain-filling duration | 3 | 0.53 | Mohammadi et al. (2008) |
| Xgwm111.2 | ACCTGATCAGATCCCCATCG | TCTGTAGGCTCTCTCCGACTG | 2B | HIS Single kernel | 5 | 0.68 | Mason et al. (2010) |
| Xgwm294 | GCAGAGTGATCAATGCCAGA | GGATTGGAGTTAAGAGAGAACCG | 2A | HIS Single kernel | 5 | 0.69 | Mason et al. (2010) |
| Xgwm356 | CCAATCAGCCTGCAACAAC | AGCGTTCTTGGGAATTAGAGA | 2A | HIS Single kernel | 5 | 0.69 | Mason et al. (2010) |
| Xwmc407 | CATATTTCCAAATCCCCAACTC | GGTAATTCTAGGCTGACATATGCTC | 2A | HIS Single kernel | 3 | 0.19 | Mason et al. (2010) |
| Xbarc137 | CCAGCCCCTCTACACATTTT | GGCCCATTTCCCACTTTCCA | 1B | Grain-filling duration | 11 | 0.63 | Mason et al. (2010) |
| Xcfd43 | CCAAAAACATGGTTAAAGGGG | AACAAAAGTCGGTGCAGTCC | 2D | Grain-filling duration | 5 | 0.67 | Mason et al. (2010) |
| Xcfa2129 | ATCGCTCACTCACTATCGGG | GTTGCACGACCTACAAAGCA | 1A | HIS kernel number of spike | 4 | 0.66 | Mason et al. (2010) |
| WMC527 | GCTACAGAAAACCGGAGCCTAT | ACCCAAGATTGGTGGCAGAA | 3B | HIS Single kernel | 6 | 0.48 | Mason et al. (2010) |
재료 및 방법
식물재료 및 Genomic DNA 추출
식물재료는 금강, 조경, 백중, 고소, 올, 올그루, 조품 등 7개 품종과 CIMMYT 터키 농업연구소로부터 분양 받은 23계통 등 31개의 밀을 사용하였다(Table 1). 터키 농업연 구소에서 분양 받은 23계통은 내고온성 평가 결과 고온에 저항성을 갖는 품종과 계통으로서 본 연구에 이용되었다. Genomic DNA는 Genomic DNA extraction Kit for plant (SolGent, Korea)를 이용하여 추출하였다.
Table 1.
Information of 31 in bred lines and wheat cultivars used in this study.
PCR 및 다형성 분석
기존 연구에서 보고된 14개 SSR 마커를 사용하여 품종 및 계통 31개를 PCR 분석하였다(Table 1, Table 2). PCR은 전체 60 ng의 DNA를 주형으로 하여, 프라이머 0.5 μmol, dNTP 0.2 mM, Taq DNA polymerase 1 unit (GenetBio., Korea)의 조성으로 수행되었다. PCR은 터치다운 방법을 이 용하여 수행되었으며, 조건은 94℃에서 30초, 65℃-55℃ (-0.7℃/cycle); 30초, 72℃에서 1분으로 13회 한 후 94℃; 30초, 58℃, 30초, 72℃; 1분으로 35회 반복하였고, QIAxcel 자동 전기영동장치(QIAGEN, USA)를 이용하여 전기영동 한 후 다형성을 확인하였다. Polymorpism information content (PIC)는 PIC = 1-Σni = 1pi2로 계산하였다(Liu, 1998).
연관관계 분석
SSR 마커 다형성 분석을 통해 확인된 밴드 중 50-400 bp 사이의 DNA 밴드를 데이터화 하였다. PCR을 통해 증폭된 밴드와 증폭되지 않은 밴드를 각각 ‘1’과 ‘0’으로 데이터화 하여 NTSYS-pc 2.0 프로그램을 이용하여 통계적 분석을 수행하였다(Rohlf, 1998). 31개 유전자형의 유전적 연관관 계는 unweighted pair-group method on arithmetic averages (UPGMA)로 나타냈다.
결과 및 고찰
SSR 마커 다형성 분석
본 연구에 사용된 14개 SSR 마커는 밀의 2A, 1B, 2D 등 서로 다른 염색체상에 존재하는 것으로서 밀 종자의 수, 무 게, 개화 후 성숙단계 등 생산성과 관련된 QTL과 가깝게 연관되어 있는 것으로 보고되었다(Sadat et al., 2013). 기존 의 많은 QTL 연구를 통해 6배체 보통밀과 듀럼밀의 전체 수량구성요소와 건조스트레스 저항성 등이 함께 평가되었 지만, 단일 생산성구성요소에 대한 유전자지도 작성 등의 접근 방법은 독자적인 것으로서 내고온성 등과 같은 하나의 형질과 관련 있는 QTL 분석으로서 시도되었다(Mason et al., 2010). 고온은 특히 개화 후 10~15일에서 완전 성숙이 되는 단계에 영향을 미치기 때문에 종자의 수와 무게의 감 소 등에 영향을 미친다(Houghton et al., 2001). 종자의 성 숙 기간의 고온은 또한 종자와 최종적으로 밀가루 품질의 저하를 야기시키는 원인 중 하나로 작용한다(Bhullar and Jenner, 1985). 본 연구에 사용된 SSR 마커는 대부분 종자 의 성숙단계와 종자 수 등의 QTL과 연관되어 있다(Table 2). 14개 마커를 국내에서 육성된 밀 품종 7개, 터키에서 분 양 받은 내고온성 유전자원 23개와 Chinese spring 등 31개 밀에 적용하여 유전자형을 비교하였다. 터키로부터 받은 품 종 및 계통은 내고온성 평가에서 고온 및 가뭄 스트레스에 저항성을 갖는 것으로 나타났다. 주요 국산밀 품종은 국내 기후에 맞게 육성된 것으로서 내고온성 특성 분석이 현재 되어있지 않다. 그러나 터키 자원과의 유전자형 분석을 통 해 내고온성 특성을 어느 정도 유추할 수 있을 것으로 생각 되고, 내고온성에 대한 국내 연구가 진행될 것이다. 31개 밀 을 분석한 결과 전체 86개의 대립유전자가 확인되었다(Fig. 1). 각각 마커의 다형성 분석은 50~400 bp 이내의 밴드를 데이터로 사용하였고, 3~16개의 대립유전자가 확인되었다. 가장 많은 대립유전자 수가 확인된 마커는 Gwm291로서 16개가 확인되었고, 가장 적은 수의 대립유전자는 Xwmc407 과 Gwm484에서 3개가 확인되었다(Table 2). Gwm291 마 커는 50~300 bp 범위에서 16개의 가장 많은 다형성 밴드가 확인되었고 PIC는 0.83이었다. Gwm11, Xcfd43, Xgwm294, Xgwm111.2의 다형성 밴드 수는 50~400 bp 범위 내에서 각각 4개, 5개, 5개, 5개로 평균 대립유전자 수와 비슷하게 나타났고, 이들의 평균 PIC는 0.66이었다(Fig. 1). Fig. 1에 서 보는 것과 같이 QIAxcel 자동 전기영동장치는 일반 아 가로스젤에서 확인하는 것보다 세밀하게 차이를 볼 수 있기 때문에 실제 대립유전자 수가 아가로스에서 확인하는 것보 다 많고, 세밀하게 분석할 수 있었다(Fig. 1). SSR 마커는 염색체상의 한 유전자좌에서도 많은 대립유전자로서 나타 날 수 있다(Naghavi et al., 2007). SSR 유전자 좌에 속해있 는 대립유전자 다양성이 많은 연구를 통해 보고되었다 (McCouch et al., 2001). 평균 6.14개의 대립유전자가 본 연 구를 통해 확인되었으며, 이것은 기존의 연구에서 보고된 5.4개(Sadat et al., 2013)와 5.2개(Zeb et al., 2009), 3.2개 (Salem et al., 2008)등과 다르게 나타났다. 14개 SSR 마커 의 평균 PIC는 0.64로 나타났으며, Xwmc407이 0.19로 가장 낮게 나타났고, Gwm293과 Gwm291이 0.83으로 가장 높게 나타났다. Xwmc407의 대립유전자 수가 3개로 적은 것에 비해 다형성 밴드가 많았기 때문으로 생각된다.

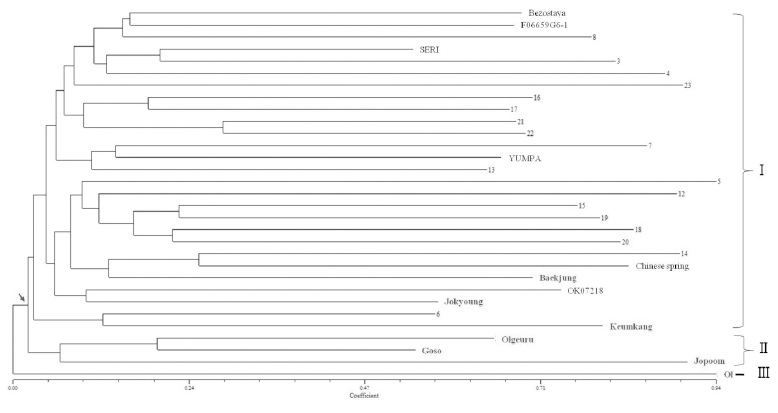
계통학적 분석
계통학적 분석결과 크게 세 개의 그룹으로 형성되었다 (Fig. 2). 국산 품종 중 올이 아웃그룹(III)으로 형성되었고 올그루, 고소, 조품, 등 국산 품종은 터키 자원과는 다르게 두 번째(II) 그룹에서 나타났다. 내고온성과 관련된 터키 자 원과 금강, 조경, 백중, Chinese spring은 첫 번째(I) 그룹으 로 형성되었다(Fig. 2). 국산밀 품종 중 올, 조품, 올그루 등 세 품종은 숙기가 빠른 품종으로서 올은 1966년 영남농업 시험장에서 농림72호와 농림12호를 교배하여 육성된 품종 으로 출수기와 성숙기가 그루밀보다 2일 빠른 것으로 보고 되어 있다(농촌진흥청, 2011). 조품은 1990년 작물시험장에 서 관동75호의 교잡종(SW88416-B-0)과 CIMMYT에서 도 입된 품종의 F1과의 교배를 통해 육성된 품종으로 또한 출 수기과 성숙기가 그루보다 5~10일로 매우 빠르고 올그루 또한 그루보다 2~5일 빠른 것으로 보고되어 있다(농촌진흥 청, 2011). 금강은 빵용, 고소는 과자용으로 용도별로 육성 된 품종이다(Nam et al., 1998). 조경과 백중은 각각 2004년 과 2007년에 육성된 품종으로 출수기와 성숙기가 우리보다 3~4일 빠른 것으로 보고되어 있다. 이들 품종은 모두 숙기 가 단축된 품종으로서 그루보다 출수기와 성숙기가 평균 3~6일 정도 빠른 것으로 보고되어 있다. 그러나 올, 올그루, 조품, 고소는 파성이 II로서 춘파형, 금강, 백중, 조경은 파 성이 III형으로 양절형으로 보고되어 있다. 춘파형인 올, 올 그루, 조품, 고소 등 4개 품종은 터키에서 분양된 내고온성 자원과 서로 다르게 그룹화 되어 있고, 금강, 조경, 백중 등 3개 품종은 내고온성 자원과 보다 가깝게 연관되어 있는 것 으로 나타났다. 이 결과를 통해 밀의 내고온성은 출수기와 성숙기 등의 특성보다 파성과 매우 밀접하게 연관되어 있는 것으로 추측된다.
적 요
밀의 등숙기간, 종자 수, 종자무게와 관련된 SSR 마커 14개를 이용하여 터키에서 분양받은 내고온성 유전자원 23개와 국산밀 품종 7개, Chinese spring 1개 등 31개를 분석한 결과, 전체 86개의 대립유전자(평균 6.14개)가 확 인되었다. 평균 PIC 값은 0.64로 나타났다. 다형성 분석을 통한 마커 데이터를 이용하여 계통 분석을 한 결과 크게 세 그룹으로 형성되었다. 올밀이 가장 바깥 그룹으로 형성 되었고 올그루, 고소, 조품 등이 터키자원과 다르게 단일 그룹으로 형성되었다. 금강, 조경, 백중 등 세 품종은 터키 자원들과 같은 그룹으로 나타났지만 밀접하게 연관되어 있지는 않은 것으로 나타났다. 국내 품종 중 올, 올그루, 조품, 고소 등 4개와 금강, 조경, 백중 등 3개가 서로 다른 그룹으로 형성되었다. 두 그룹의 차이는 파성 II와 III의 차 이로 구별되었다.
