

벼의 수발아는 황숙기에서 완숙기 사이에 발생하기 쉬우며 (Park & Park, 1984) 우리나라와 같은 온대지역에서는 등숙 기의 강우나 태풍으로 인해 주로 피해가 발생한다(Suh & Kim, 1994). 벼 수발아가 발생하면 완전립률과 쌀 외관품질 이 저하되며 현미천립중이 감소하여 수량도 저하된다(Kim et al., 2008; Kim & Lee, 1996; Lee et al., 1985; Oh et al. 1987). 벼의 수발아는 재배 시기, 수확 시기, 일장, 온도의 영향에 따라 차이가 있으며(Cho et al., 1987), 배수가 불량 한 곡간답이나 다비 조건에서 발생이 심하다고 한다(Park & Park, 1984). 수발아성은 유전적 차이도 커 인디카 품종이 자포니카 품종보다 수발아 저항성이 크다(Ju et al., 2000; Kim & Lee, 1996; Kim, 1995; Suh & Kim, 1994). 품종의 조만성에 따라서는 조생종이 중만생종보다 수발아성이 크 다는 보고도 있으며(Park & Park, 1984) 조생종에도 수발아 성이 낮은 품종이 있다는 보고(Park & Kim, 2009)도 있어 이견이 있다. 종자의 휴면성이 강한 품종은 수발아 저항성도 큰 것으로 알려져 있으며(Bewley, 1997; Gianinetti & Vernieri, 2007), flavonoid 색소를 가진 적색미도 수발아에 강한 것으 로 알려졌다(Gu et al. 2011).
밀, 라이밀의 수발아 종자는 종자 활력이 낮아 파종기의 발아율에 영향을 미친다고 한다(Elias & Copeland, 1991; Barnard & Calitz, 2011; Mos, 2003). 또 수발아립은 수확 후 저장 중 미생물에 감염되기도 쉽다(Castor & Frederiken, 1977; Mares, 1989). 벼는 수발아 저항성이 클수록 종자 저 장 수명과의 상관이 높다고 하며(Lee et al., 2006), 수발아 종자의 싹이 길수록 성묘율이 저하된다는 보고가 있다(NICS, 2006). 최근 들어 채종용 포장의 벼 보급종자에서 발아가 불 균일한 경우가 증가하고 있는데 이러한 품종들은 대부분 정 조의 정립율은 우수하나 현미 피해립율과 수발아립률의 차 이가 큰 경우가 많았다. 그러나 벼 수발아는 수량과 미질에 미치는 영향에 대한 보고가 대부분으로(Kim et al., 2008; Kim & Lee, 1996; Lee et al. 1985; Oh et al. 1987) 저장수 명과 종자 발아에 미치는 영향에 관한 자료가 매우 부족하다. 벼 종자는 현미를 왕겨가 싸고 있기 때문에 수발아가 발생했 더라도 정조 상태에서 수발아 피해정도를 판별하기 쉽지 않 고 판별 기준이 없는 실정이다. 따라서 본 시험에서는 벼 종 자의 수발아립률 차이가 파종기의 출아율에 미치는 영향과, 발아정도에 따른 수발아립의 형태와 재발아율을 검토하였다.
재료 및 방법
시험재료의 현미외관품위 차이
시험재료는 2013년 농업기술실용화재단의 철원과 김제 채종포 2 곳(김제-1, 2)에서 수확한 호품벼 중 완전립률과 수발아율의 차이가 큰 두 시료를 사용하였다. 시료는 수확 후 정선작업을 거친 후 동일한 조건에서 이듬해 파종기까지 저장하였다. 현미 품위는 곡물검사기(Cervitec 1625 Grain inspector, Foss co.)로 조사한 후 농업과학기술 연구조사분석 기준(2012)에 준하여 다시 육안으로 재분류하였다.
3군데 포장의 호품 시료 중 철원과 김제2 호품은 완전미 율이 88.6%, 87.1%로 차이가 거의 없었으나 김제-1 호품은 77.2%로 두 시료에 비해 완전미율이 10% 정도 낮았다(Table 1). 김제-1은 수발아립률이 8.4%로 철원(1.4%), 김제2(3.7%) 에 비해 높았다. 그러나 3가지 시료 모두 정조의 외관상 수 발아 흔적을 찾기가 쉽지 않았다. 3가지 호품의 현미외관품 위는 지역 간 차이를 보이지 않았고 동일 지역 내에서 오히 려 차이가 컸다.
Table 1.
Grain quality of hulled rice of Hopum from three harvest regions.
| Variety | Harvest region | Head (%) | Imperfect (%) | PHS* | |||
|---|---|---|---|---|---|---|---|
| Immature | Damaged | Dead | Total | ||||
| Hopum | Cheorwon | 88.6 a | 6.9 b | 3.4 c | 1.0 b | 11.4 b | 1.4 c |
| Hopum | Gimje-1 | 77.2 b | 8.3 a | 11.3 a | 3.2 a | 22.8 a | 8.4 a |
| Hopum | Gimje-2 | 87.1 a | 4.0 c | 6.7 b | 2.2 b | 12.9 b | 3.7 b |
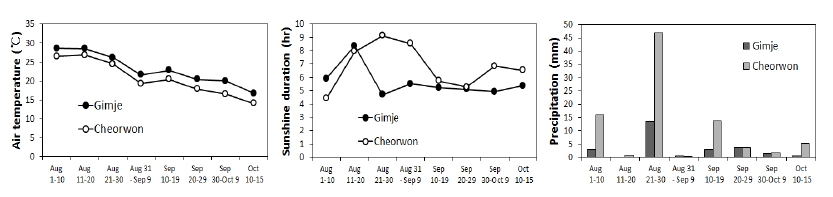
채종지 등숙기 기상 분석에는 기상청 자료를 사용하였는 데, 김제는 기상관측소가 없어 가까운 익산의 기상자료를 사용하였다. 수확지의 등숙기(8.11~10.15) 평균기온은 익산 (김제)이 철원보다 2.4도 높았으며, 순별 일조시수와 강수량 은 8월 하순에 두 지역의 차이가 컸는데 철원이 일조시수와 강수량 모두 높았다. 9월 상순까지 일조시수가 철원이 익산 보다 높았으나 이후 강수량과 일조시수는 큰 차이가 없었다 (Fig. 1). 호품 출수기는 8월 20일 경으로 9월 중순 이후의 기상이 수발아에 영향을 미칠 가능성이 크지만 9월 중순 이 후 두 지역의 강수량이나 일조시수의 차이가 없어 기상에 의한 영향은 거의 없었을 것으로 생각되었다. Park & Park (1984)은 배수와 통풍이 불량하고 다비할 경우 수발아 발생 률이 높다고 하였는데, 본 시험에서 동일지역에서 표준재배 법을 따른 채종포에서 이와 같이 현미 품위 차이가 큰 것은 토질의 비옥도나 물관리에 의한 생육 차이의 영향 때문일 것으로 생각된다. 따라서 3가지 호품 시료의 현미 품위 차 이는 지역 간 등숙 기상 차이가 아니라 포장 및 재배관리로 인한 군락 내 환경의 차이로 발생한 것으로 생각된다.
발아특성, 출아율, 종자활력
출아율 조사는 수확 후 6개월 뒤인 4월 하순에 실시하였 다. 종자소독(procloraz와 fludioxonyl 2000배 혼용액, 30°C, 24시간) 후 침종하여 균일하게 싹이 1~2 mm로 틀 때 육묘 상자 당 130 g씩 5반복으로 파종하여 30°C의 육묘기에서 2 일간 출아시키고 육묘온실에서 8일간 생육시킨 후 출아율과 모 생육을 조사하였다. 발아시험은 페트리디쉬(10×4 cm)에 여과지 2장을 깔고 각각 100립씩 4반복으로 균일하게 종자 를 치상하고 여과지를 덮은 후 증류수 10 ml을 분주한 다음 25°C에서 처리하였다. 발아율(germination percentage)은 치 상 후 10일의 최종발아율로, 발아속도는(germination speed) 는 치상 후 일별 발아립수의 총 합계로 표시하였다. 종자활 력 검정은 품종별로 정조 50립씩을 20°C에서 18시간 물을 흡수시킨 다음, 배가 노출되도록 종단면으로 절단하고 1% 2,3,5-triphenyl tetrazolium chloride (TTC), pH 7.0 용액에 서 30°C, 1시간 염색한 후 증류수로 2~3번 헹구어 내고 배 단면의 착색정도로 판단하였다(AOSA, 2007).
수발아립의 외관 형태와 발아율
현미 품위별로 완전립, 미숙립, 피해립을 페트리디쉬에 50립씩 3반복으로 치상하고 25°C에 처리하여 7일 뒤 발아 율을 조사하였다. 현미 수발아립은 완전립에 비해 배가 돌 출되었으며 검은색으로 변색되었고 배유가 투명하지 않고 흰색 또는 갈색으로 변색된 정도로 판별하였다(Fig. 2). 현 미 형태 변화가 진행된 정도를 Fig. 2와 같이 분류하고, 수 발아립을 2단계(PHS I, II)로 구분하여 발아율을 조사하였 다. 수발아립의 표면 살균은 3% 차아염소산나트륨액에 20 분간 처리하고 증류수로 3회 세척 후 페트리디쉬에 치상하 였다.
종자 발아 처리 및 퇴화촉진처리
수발아립의 형태 변화와 재발아력의 변화를 추정하기 위 해 싹 틔워 건조시킨 종자의 형태와 재발아력을 퇴화 촉진 처리하여 비교하였다. 호품벼를 25°C의 수돗물에 침종하여 매일 물을 갈아주다가 싹이 트기 시작하면 싹 길이 1 mm미 만, 1~2 mm, 2 mm로 분류하여 실내에서 10일간 건조시켰 다. 건조시킨 정조를 페트리디쉬에 BP (BP: between paper) 법으로 50립씩 3반복으로 치상하고 25°C에 처리하여 7일 뒤 발아율을 조사하였다. 발아립의 퇴화촉진처리(AA: accelerated aging)는 플라스틱 밀폐용기(12×12×6 cm)에 증류수 40 ml 을 넣고 3 cm 높이의 스테인리스 트레이를 넣고 트레이 위 에 종자 50립씩을 단층으로 고루 펼친 후 뚜껑을 닫아 밀폐 하고 45°C에 24시간 처리하였다.
수발아립과 발아립의 형태는 실체현미경(SZX16, Olympus co.)으로 관찰하였다. 데이터 통계분석은 SPSS statistics (Ver. 19)를 사용하였으며, 처리간 평균값을 던컨다중범위검정으 로 5% 유의수준에서 비교하였다.
결과 및 고찰
수발아립률 차이에 따른 발아특성, 출아율, 종자 활력
수발아율의 차이가 출아율과 발아율에 미치는 영향을 알 아보기 위해 현미 품위가 차이나는 두 지역(철원, 김제-1)의 호품벼를 비교하였다. 두 시료의 수발아립률은 철원 1.4%, 김제-1 8.4%였으며 완전립률은 각각 88%, 77%였다(Table 1). 발아율은 철원은 95.7%로 김제-1보다 발아율이 30% 이 상 높았으며 발아속도도 빨랐다(Table 2). 종자활력은 철원 96.3%, 김제-1 66.0%로 나타나 발아율과 유사한 결과를 보 였다. 상토에 파종하여 정상적으로 입모한 출아율은 철원 호품이 93%, 김제-1 호품이 61%로 32% 차이를 보였으나 (Table 2), 정상모의 엽수, 초장, 근장은 각각 1.16매, 5.27 cm, 3.64 cm로 모두 동일하여 출아율은 차이가 있어도 정 상모의 생육 차이는 없었음을 알 수 있었다.
Table 2.
Germination percentage, germination speed, seedling emergence rate and seed viability of Hopum in two regions.
철원 호품은 발아율과 출아율이 완전립률보다 높았으나, 김제-1 호품은 완전립률보다 오히려 낮았다. Barnard (2011) 는 밀의 경우 수발아율이 낮은 경우 발아율 차이가 크지 않 지만 수발아율이 높을수록 발아율의 저하가 더 크다고 하였 는데, 본 결과에서처럼 벼에서도 유사한 경향을 나타내었 다. NICS (2006)의 보고에서도 벼의 수발아율이 높으면 발 아율과 성묘율이 떨어진다고 하였다. 수발아율이 높은 시료 가 발아율과 출아율이 더 크게 감소한 결과는 수확 당시 수 발아율이 높으면 발아정도의 변이가 크고 저장기간 동안 노 화가 일어나기 때문이라는 Elias & Copeland (1991)의 보 고와 유사하다. 수발아 보리 종자도 저장하는 동안 활력이 상대적으로 빨리 저하된다고 한다(Bason et al., 1991; Moor, 1987). 따라서 수발아립률이 높고 피해립률이 높은 종자는 수확 당시보다 저장 기간 동안 종자 퇴화가 빨리 일 어나기 때문에 수발아립률 차이보다 파종기의 발아율과 출 아율이 더욱 낮은 것으로 생각된다.
수발아립의 변색 정도가 발아율에 미치는 영향
전체 발아율의 저하가 수발아립에 의한 영향인지를 알아 보기 위해 현미품위별로 발아율을 조사하고 다시 수발아립 만을 분리하여 발아율을 조사하였다. 철원과 김제-1시료를 완전립, 미숙립, 피해립으로 분리하여 발아율을 조사하였다 (Table 3). 완전립의 발아율은 두 시료 모두 99% 이상이었 다. 그러나 불완전립의 발아율은 두 시료의 차이가 컸는데, 미숙립의 발아율은 철원(71%)보다 김제-1(34%)가 두 배 정 도 높았으며, 손상립의 발아율도 철원이 김제-1보다 2배 이 상 높았다. 이때 미숙립과 손상립에 포함된 수발아립에서 세균성 부패와 곰팡이 발생이 심했는데 철원 호품보다 김제 -1 호품의 균 발생이 심했다.
Table 3.
Germination percentage of head, immature and damaged grains of rice.
| Variety | Harvest region | Germination percentage (%) | ||
|---|---|---|---|---|
| Head | Immature | Damaged | ||
| Hopum | Cheorwon | 99.7 ± 0.5 | 70.7 ± 6.8 | 53.3 ± 15.2 |
| Hopum | Gimje-1 | 99.7 ± 0.5 | 34.0 ± 8.5 | 18.7 ± 10.5 |
김제-1 시료는 철원 시료보다 수발아율이 높을 뿐만 아니 라 수발아립의 변색정도가 심했다. 따라서 수발아립을 배와 배유의 변색 정도에 따라 2 단계로 나누어(Fig. 2) 발아율을 살펴보았다. 수발아립 I은 배가 완전립보다 돌출하고 배유 의 1/3이 흰색으로 변색된 정도를 기준으로 구분하였고(Fig. 2. b, c), 수발아립 II는 I보다 배가 약간 더 돌출하고 배유가 1/2 정도 변색된 정도를 기준으로 구분하여(Fig. 2. d, e) 발 아율을 조사하였다. Kim & Lee (1996)는 수발아 처리기간 이 길어질수록 완전미율이 떨어진다고 하였는데, Kim et al. (2008)은 이는 수발아한 배와 배유가 착색되기 때문에 불완전미가 증가하였기 때문이라고 하였다. 따라서 수발아 립 I과 II의 형태 차이는 수발아 발생 기간에 따른 차이로 생각되며 변색 정도가 심할수록 수발아 발생 기간이 길었을 것으로 생각된다.

수발아 I과 II의 발아율은 철원 호품은 27.3%, 12.7%였 고 김제-1은 9.3%, 0%로 김제-1이 철원보다 발아율이 낮았 고 곰팡이 발생도 심했다(Table 4, Fig. 3). 발아속도가 느린 종자가 주변의 균에 감염되어 미발아 할 가능성을 배제하기 위해 현미 표면을 차아염소산나트륨으로 소독한 후 발아율 을 조사하였다. 소독 후 수발아립의 발아율은 철원은 수발 아립 I과 II가 각 각 42%, 26%로 미소독 처리에 비해 발아 율이 증가하였으나, 김제-1은 두 수발아립이 모두 8%로 수 발아립 II의 발아율이 증가하기는 하였으나 발아 후 정상적 으로 생육하지 못하고 부패하였다. 이 결과로 보아 수발아 율이 낮은 철원 호품은 종자 감염이 심하지 않아 수발아립 에서도 일부가 발아가 되며 미숙립의 발아율도 높아 전체 발아율이 완전립보다 높았던 것으로 생각된다. 반면 상대적 으로 수발아율이 높았던 김제-1 호품은 대부분의 수발아립 에 균사가 배유조직까지 침투하였기 때문에 표면의 균사가 살균이 되지 않았고, 발아속도가 느린 주변의 종자까지 영 향을 미쳤을 것으로 생각된다. 수수와 밀의 수발아 종자는 미생물의 감염이 쉬워 종자 부패의 원인이 된다고 하였는데 (Castor & Frederiken, 1977; Mares, 1989), 본 시험에서 발 생한 곰팡이 포자 관찰 결과 특정 벼 병균은 아니었으며 일 반적인 부생균으로 판단되어 수발아가 심할수록 포장에서 나 저장 중에 균의 감염이 많이 발생했을 것으로 생각되었 다. Kim et al. (2008)은 수발아가 진전될수록 불완전립률이 증가하는 것은 수발아립의 조직의 변화가 심해지기 때문으 로 추측하였는데, 본 실험에서 수발아율이 높은 시료가 배 와 배유의 모양과 착색이 심해진 것을 확인할 수 있었다. 이 는 수발아가 진전되면서 배와 배유의 양분소실이 커지고 미 생물에 의한 감염에 따라 착색이 심해졌기 때문으로 생각된다.
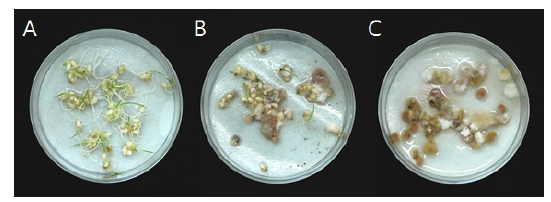
Fig. 3.
Germination of head(A), PHS-I(B) and PHS-II(C) grains of Hopum in Gimje-1. Head grains were vigorously germinated but pre-harvest sprouted grains were rotted and infected by fungi.Table 4.
Germination percentage of pre-harvest sprouted(PHS) grains.
| Variety | Harvest region | Unsterilized | Sterilized | ||
|---|---|---|---|---|---|
| PHS-I | PHS-II | PHS-I | PHS-II | ||
| Hopum | Cheorwon | 27.3 ± 2.5 | 12.7 ± 5.2 | 42.0 ± 3.3 | 26.0 ± 3.3 |
| Hopum | Gimje-1 | 9.3 ± 1.9 | 0.0 | 8.0 ± 2.8 | 8.0 ± 1.6 |
수발아립과 발아 종자의 형태 비교
위의 결과로 수발아립의 형태변화가 심할수록 발아율이 크게 떨어진 것을 확인하였다. NICS (2006)의 보고에 따르 면 수발아 종자를 15일간 건조 후 재발아율을 보면 싹이 길 수록 발아율과 성묘율이 떨어진다고 하였으며, Oh et al. (1987)는 수발아 유아 길이가 1.6 mm이상이면 완전립률이 20% 감소한다고 하였다. 그러나 포장에서 경미한 수발아가 발생하였더라도 수확과정을 거치면서 기계적 손상으로 싹 이 부러지기 때문에 싹이 얼마나 자랐었는지를 알기 힘들 다. 따라서 현미의 배와 배유의 형태로 수발아 정도를 추정 하는 것이 보다 정확한 수발아율을 확인하는 방법이지만 이 에 관한 형태적 보고가 거의 없었다. 따라서 본 시험에서는 수발아 진행정도에 따른 형태를 추정하기 위해 실험실에서 종자를 발아시켜 배와 배유의 변색 정도를 관찰하였다. 종 자를 침종하여 싹을 틔운 후 싹 길이별로 건조시켜, 정조와 현미의 외관형태를 관찰하였다(Fig. 4). 발아 후 건조시킨 정조는 모두 배 위치의 왕겨가 파열되어 있었으나 싹 1 mm 미만은 왕겨 밖으로 싹이 전혀 보이지 않았고, 싹이 3 mm 이상이면 건조 상태에서도 왕겨 밖으로 싹이 보였지만, 싹 이 부러질 경우 싹튼 정도를 구분할 수 없었다. 그러나 왕겨 를 벗겨내고 현미를 관찰해보면 싹이 길수록 배의 돌출 정 도가 심하였고 싹이 3 mm이상이면 배유의 1/2정도가 변색 되었다(Fig. 4). 싹을 틔운 후 건조시킨 종자의 배를 TTC용 액으로 염색하여 활력 부위를 보면 싹 2 mm 이내의 건조 종자는 유관속, 배반, 1, 2엽과 근초 부위가 착색이 되었으 나(Fig. 4. a, b, c), 싹이 3 mm이상은 유관속을 제외한 부분 은 거의 착색이 되지 않았다(Fig. 4. d, e). 수발아립(Fig. 2) 과 발아종자(Fig. 4)의 형태를 비교하면 Fig. 2의 b, c에 해 당하는 수발아립은 Fig. 4의 A (싹 1 mm >), B (싹 1~2 mm) 의 형태와 비슷하였으며, Fig. 2의 d, e는 Fig. 4의 C (싹 3 mm), D (4 mm)와 배유의 변색정도가 유사하였다. 따라서 수발아립의 배유가 1/2이상 변색된 정도이면(Fig. 2의 d, e) 싹이 3 mm이상 발아하였을 것으로 생각된다.

Fig. 4.
Morphological appearance of sprouted grains of unhulled and hulled rice according to shoot length. Rice seeds were sprouted by soaking in water and dried for 10 days at room temperature (A, B, C and D represent 1 mm >, 1~2 mm, 3 mm and 4 mm of shoot length). Tetrazolium stained seeds of a, b, c, d and e are 1 mm >, 1, 2, 3, and 4 mm of shoot length, respectively.발아종자의 재발아율과 퇴화촉진처리 효과
실험실에서 발아시킨 종자를 건조 직후와 퇴화촉진처리 후 재발아율을 조사하였다. 발아종자의 건조 후 재발아율은 싹 1 mm 미만, 2 mm, 3 mm에서 각 각 98.7%, 92.7%, 43%로 나타나 싹이 3 mm이상 발아하면 재발아율이 크게 저하됨을 알 수 있었다(Fig. 5). NICS (2006)는 수발아 처리 직후 15일간 건조시킨 종자의 재발아율이 4 mm부터 크게 저하되었다고 하였는데 본 시험에서 발아시킨 종자 건조 후 재발아율이 3 mm이상에서 크게 저하되는 점과 유사하였다.
퇴화촉진처리한 발아종자의 재발아율은 싹 1 mm 미만, 2 mm, 3 mm가 각 각 80%, 25%, 3%로 나타나 무처리에 비해 각 각 19%, 68%, 40% 감소한 것으로 나타났다(Fig. 5). 특히 싹 2 mm이상부터 재발아율이 크게 감소하였다. Mos (2003)는 퇴화촉진처리에 의한 발아율 저하 경향이 수 발아립의 저장기간과 품종에 따른 발아율저하와 매우 상관 이 크기 때문에 수발아립의 저장성을 평가하는데 퇴화촉진 처리가 매우 유용하다고 하였다. 따라서 본 실험에서도 퇴 화처리에 의한 발아종자의 재발아율 경향은 수발아 종자의 저장 후 발아율과 상관이 클 것으로 생각되며, 이 결과를 바 탕으로 수발아 종자의 싹이 2~3 mm이상 발아할 경우 파종 기 발아율이 저하될 가능성이 클 것으로 생각된다.
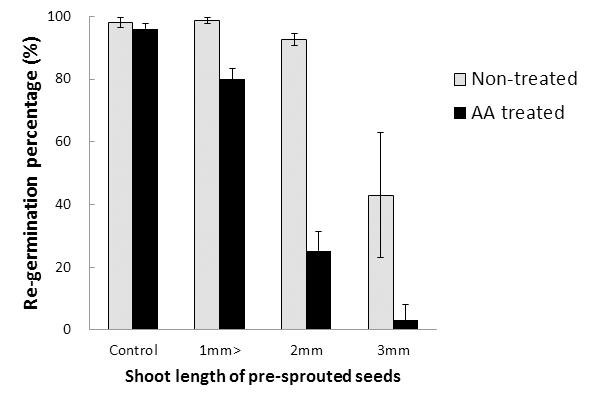
Fig. 5.
Re-germination percentage after AA treatment in presprouted seeds according to shoot length. AA (accelerated ageing) condition : 45°C, 98% RH for 24hrs.종자의 발아력이 감소하는 요인은 단기간의 요인으로는 수 발아와(Coolbear, 1995) 종자감염에 의한 영향(McGee, 2000) 이 크고 장기간에 걸친 요인은 저장기간동안 온도와 습도에 의한 영향으로 유전자, 세포막, 효소, 단백질의 손상과 호흡 활동의 변화로 일어난다고 한다(Coolbear, 1995; McDonald, 1999). Stahl & Steiner (1998)은 수발아율이 높으면 활력이 낮고 저장 온도와 습도가 높으면 활력이 상실되지만 두 가지 조건은 독립적인 요인으로 작용한다고 하였다. 따라서 벼에 서도 수발아 정도가 심할수록 종자활력이 저하되고 파종기 발아율을 저하시키는 요인으로 작용한다고 할 수 있다.
본 결과와 같이 수발아는 동일 품종에서도 포장 간 차이가 크게 발생할 수 있으므로 수발아 방지를 위해서는 토양별로 통풍, 배수, 시비량 등의 재배적인 측면에서 관리가 요구된다. 최근 이와 같은 국지적인 벼 수발아 발생이 문제가 되고 있으 나 국내 육성 벼 품종에 대한 수발아의 생리·생태적 연구가 미 흡한 실정으로 이에 관한 추후 연구가 필요하다고 생각된다.
적 요
생산지에 따라 벼 종자의 수발아립률 차이가 컸던 호 품벼의 출아율과 발아율을 조사한 결과, 수발아율 8% 인 시료는 수발아율이 1%인 시료에 비해 발아율과 출 아율, 종자활력이 30% 낮았다.
수발아율이 높은 시료는 수발아립의 변색 정도가 심 하고 미발아율과 부패율이 높았다. 수발아립은 배가 돌출하고, 변색된 배유와 곰팡이 감염이 관찰되었으며 배유의 1/2이상이 변색된 종자는 발아율이 매우 낮았다.
종자의 발아 진행정도에 따라 현미의 변색이 심해졌 는데, 싹이 3 mm이상일 경우 배유의 1/2정도가 변색 되었다.
발아 종자는 싹이 3 mm이상일 때부터 재발아율이 크 게 떨어졌으며 퇴화촉진처리 후 발아율의 감소가 더 컸다. 특히 퇴화촉진처리한 발아 종자는 싹이 2 mm부 터 종자활력이 크게 감소하였다.
결론적으로 수발아 발생이 심할수록 파종기 발아율과 출 아율의 감소가 크고, 수발아가 3 mm이상 발생하면 발아율 이 크게 저하될 것으로 생각된다. 또한 수발아는 동일 품종 과 동일 지역에서도 포장 간 차이가 크게 발생할 수 있으므 로 재배관리에 주의해야 한다.
