

Rapeseed(Brassica napus L.) has become one of the world's most important sources of vegetable oil due to extensive breeding and cultivation of this crop. In general, rapeseed oils contain about 65% oleic acid (C18:1), 20% linoleic acid (C18:2), and 10% linolenic acid (C18:3). A balanced fatty acid mixture of C16/C18 chain lengths is good for nutritional uses, whereas oleochemical applications prefer vegetable oils that contain a single fatty acid only (Röbbelen & Kräling, 1993). Erucic acid has traditionally been the required constituent in rapeseed oil, but rapeseed varieties with high oleic acid have recently been developed (Röbbelen & Kräling, 1993). The high level of oleic acid and corresponding high degree of oxidative stability in oilseeds is valuable for high-cost uses in the manufacture of certain cosmetics and pharmaceuticals (Röbbelen & Kräling, 1993). The development of rapeseed cultivars with high oleic and low linolenic acid, which confers good low-temperature fluidity, is highly desirable for biodiesel (Jang et al., 2010; Lee et al., 2010).
Improving the fatty acid composition and increasing the oil content in the seed are important breeding goals for B. napus (Zhao et al., 2008). Mutation breeding has become a successful tool for manipulating the composition of fatty acids in oilseeds (Bühsenschüz-Nothdurft et al., 1998; Röbbelen & Kräling, 1993; Savant & Kothekar, 2011). Ethyl methanesulfonate (EMS) has been shown to be one of the more potent chemical mutagens available (Jacobs, 1969). EMS mutagenesis has been used successfully to change the oil content and fatty acid composition of several oilseed crops including B. napus (Auld et al., 1992; Spasibionek, 2006), soybean (Patil et al., 2007; Rahman et al., 1994), sesame (Savant & Kothekar, 2011), Arabidopsis (James & Dooner, 1990), and camelina (Bühsenschüz- Nothdurft et al., 1998). In Brassica species, Rakow (1973) obtained the first low-C18:3 mutant (M57) in zero C22:1 B. napus, with a reduction in C18:3 content from 9.7% to 5.6% in the seed oil. Auld et al. (1992) screened a mutant (X-82) from 39,504 M2 seeds of B. napus, which had 6.6% polyunsaturated fatty acids (PUFA) vs. 27.4% PUFA in ‘Cascade’. In addition, several M3 and M4 lines derived from X-82 had <6% total PUFA and >88% oleic acid. Development of EMS-induced mutants for improved seedand/ or oil-quality traits has been conducted in Ethiopian mustard (Brassica carinata A. Braun) (Sheikh et al., 2009).
In Korea, no rapeseed variety has been developed using EMS-induced mutation breeding. The objective of this study was to develop rapeseed mutants with beneficially altered fatty acid composition from B. napus ‘Tamla’ variety by EMS mutagen treatment. Imbibed seeds were treated with 1% EMS and a wide range of variability was observed for all the fatty acids in M2 population. We obtained two mutants with >75% C18:1 through induced mutagenesis of B. napus ‘Tamla’ with 69% C18:1 in M3 population. This paper further describes two single-plant progenies with altered composition of C18 unsaturated fatty acids in M4 population.
MATERIALS AND METHODS
EMS treatment
Imbibed seeds of rapeseed (Brassica napus L.) ‘Tamla’ variety were treated with 1% EMS solution at room temperature for 3, 12, and 24 h. The seeds were then rinsed under water for >4 h and dried on filter paper at room temperature. The seeds were sown into soil in a greenhouse at the Bioenergy Crop Research Center, National Institute of Crop Science, Muan, South Korea in December 2009.
Isolation of mutants
Seeds of M1 plants were harvested in May 2010 and the progeny of M1 plants were sown into soil in October 2010. A total of 1,050 individual M2 plants were harvested in June 2011. About 100 M3 lines from M2 plants with high oleic acid composition were sown in October 2011. Selected M3 plants were grown and harvested individually. M4 seeds of selected M3 plants having the desired fatty acid composition were sown in October 2012. M5 seeds of selected M4 plants were analyzed for fatty acid composition. The selected plants in each generation were maintained in isolation from insect pollinators.
Fatty acid analysis
The fatty acid contents of seeds were analyzed by gas chromatography using a 7890A GC system (Agilent Technologies, CA, USA). The front inlet and detector temperatures were 200°C and 250°C, respectively. The initial column temperature was started at 140°C and was increased to 250°C at a rate of 5°C min-1. The flow rates of helium (He) and hydrogen (H) were 2 and 35 ml min-1, respectively. Fatty acids were identified by comparing the retention times against those of standards (Sigma Co.).
RESULTS AND DISCUSSION
Induction of variability in fatty acid composition
Our objective was to identify changes in fatty acid composition among Brassica napus L. ‘Tamla’ mutants after EMS treatment. 588 M3 seeds harvested from M2 mutant plants were analyzed. Fatty acid compositions differed substantially between untreated and mutant ‘Tamla’ seeds (Table 1). The content of oleic acid ranged from 76.9% to 13.5% and erucic acid ranged from 0% to 49.3%. Oleic and erucic acid contents showed more variability compared to the other fatty acids. According to variability of oleic acid content, the 588 mutants could be classified into two groups, Group I (high oleic acid and almost no erucic acid) and Group II (low oleic acid and high erucic acid) (Fig. 1(A)). In Group I mutants, variability in oleic acid content from approximately 55% to 77% was accompanied by simultaneous variability in linoleic acid and linolenic acid level. In Group II mutants, a decrease of oleic acid content from approximately 55% to 13% was accompanied by a major increase in erucic acid, from approximately zero to 49% (Fig. 1(A), (C)) and by a minor increase in eicosenoic acid (0.4% to 19%). Group I included about 480 mutants (82%), and the remaining 108 mutants were included in Group II. In rape and turnip rape, the synthesis of fatty acids proceeds in two directions from oleic acid: one towards eicosenoic erucic acid and the other towards linoleic and linolenic acid (Jönsson, 1977). The quantities of eicosenoic and erucic acid produced are influenced by the efficiency of the enzyme that regulates their synthesis. Low efficiency of the pathway from oleic towards linoleic acid means that a large amount of oleic acid is available for the synthesis of eicosenoic and erucic acid (Jönsson, 1977). Craig (1961) also reported that a low oleic acid and a high erucic acid levels were found in high erucic-acid strains of B. napus L., while the converse was observed in low erucic-acid strains. However, the fatty acid composition in Group II (low oleic acid and high erucic acid) in the present study needs to be confirmed. Overall, TR-458 and TR-544 in M2 population contained 74.7% and 76.9% oleic acid, respectively, which was an increase of nearly 5% and 7% compared to untreated B. napus L. ‘Tamla’.
Table 1.
Variation of fatty acid composition (% of total acids) in M2 mutants and untreated ‘Tamla’
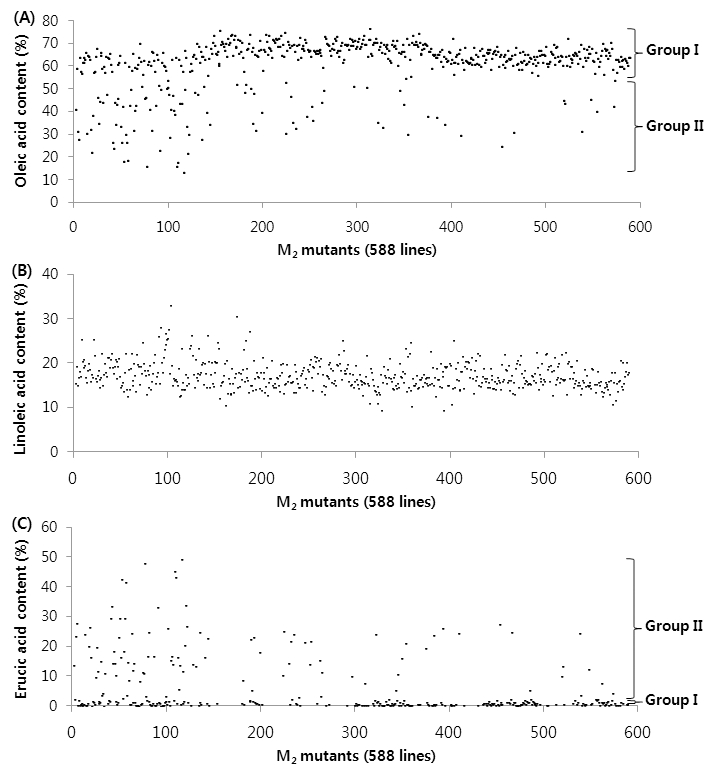
Variability of fatty acid composition in M3 mutants
To investigate the stability of oleic acid level in the TR-458 and TR-544 mutants, the M3 seeds of the mutants were grown to plants and oleic acid content in M4 seeds of the single-plant progenies was analyzed (Fig. 2). M4 seeds of TR-458-2 and TR-544-1 had ≥ 75% oleic acid content and a reduction in linoleic and linolenic acids equivalent to the increase in oleic acid (Fig. 3). TR-458-2 had increased oleic acid (75.9%) and decreased linoleic acid (12.5%) and linolenic acid (4.4%) contents; TR-544-1 had increased oleic acid (75.7%) and decreased linoleic acid (13.5%) and linolenic acid (3.3%) contents. Under same growth conditions, untreated B. napus L. ‘Tamla’ had 69.5% oleic acid, 17.5% linoleic acid, and 5.4% linolenic acid contents. Compared to the untreated ‘Tamla’ seeds, linoleic and linolenic acid contents in the mutant lines decreased by nearly 5~6% and 1~2%, respectively. Similar changes in fatty acid profile after EMS treatment have been reported in winter rapeseed (Spasibionek, 2006) and Ethiopian mustard (Velasco et al., 1997). Spasibionek (2006) selected two mutants, M-10453 and M-10464, with increased levels of oleic acid (approximately 76%) and reduced linoleic and linolenic acid contents (8.5% and 7.5%, respectively). It was suggested that mutations one or more genes controlled desaturation of oleic acid in these plants (Spasibionek, 2006). In the present study, the agronomic performance of the selected M3 plants was also evaluated. Yield components of the mutant plants were similar to those of control plants grown under the same conditions (Table 2). Seed oil content of the two mutant lines averaged 35%. Failure to achieve a substantial increase in oil content reflected a lack of selection pressure, as selection in each generation was carried out only for the desired fatty acid composition.
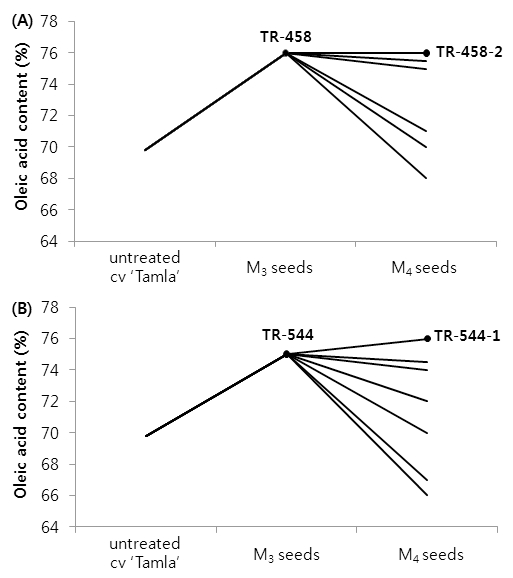
Fig. 2.
Variability in oleic acid content in M3 and M4 seeds of two single-plant progenies (TR-458 and TR-544). Mutants TR-458-2 and TR-554-1 have increased oleic acid content (>75%) in comparison with untreated cv ‘Tamla’.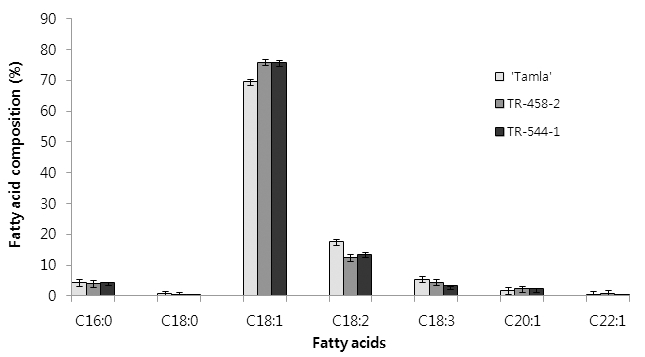
Fig. 3.
Fatty acid composition of M4 seeds of two mutants (TR-458-2, TR-544-1). The mutants showed increased oleic and decreased linoleic and linolenic acid levels in comparison with untreated cv ‘Tamla’.Table 2.
Yield components of selected M3 mutant plants. The oil content is expressed in terms of percentage.
Variability of fatty acid composition in M4 mutants
Two single-plant progenies, TR-458-2 (95 plants) and TR-544-1(118 plants), were further examined to investigate the stability of fatty acid composition in the M5 seeds. The variability in oleic, linoleic, and linolenic acid content in individual M5 seeds is shown in Fig. 4. In general, the respective positive and negative shifts in oleic and linoleic acid contents were still observed in M5 seeds. The oleic acid content in TR-544-1 and TR-458-2 progenies was 61~75% and 58~75%, compared to 68~70% in ‘Tamla’; linoleic acid content was 11~22% and 11~24%, respectively, compared to 13~15% in ‘Tamla’; and linolenic acid content in the respective progenies was 4~8% and 3~7%, compared to 4~6% in ‘Tamla’. The accumulation or reduction of oleic acid was accompanied by a simultaneous decrease or increase in linoleic and linolenic acid content (Fig. 4). M5 seeds of the two single-plant progenies were not stable and the variability in C18 unsaturated fatty acid contents was wider than in the control, indicating that these effects were the result of mutation events. In soybean oil, EMS treatment was effective in increasing the variability of fatty acid content (Patil et al., 2007). The variability was skewed towards high levels of oleic acid (35–42%) and low levels of linolenic acid (3.8–5.0%). High-oleic variants were stable in the M3 and M4 generations (Patil et al., 2007).
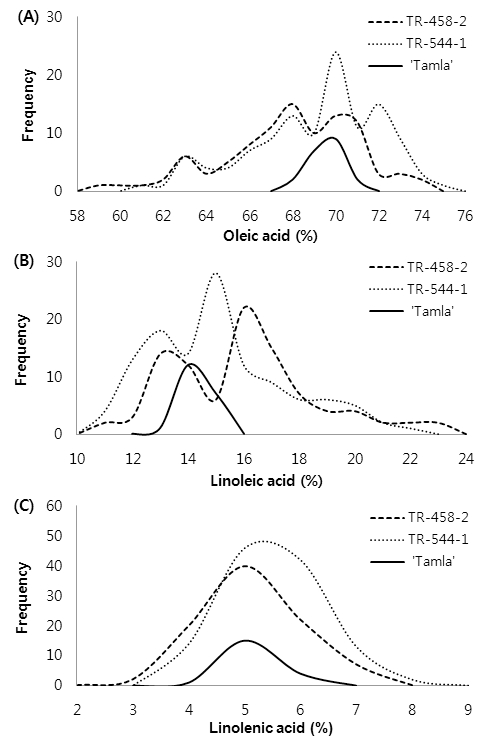
Fig. 4.
Frequency histogram showing variability in oleic, linoleic, and linolenic acid content in individual M5 seeds. Values represent 95 lines (TR-458-2 progeny), 118 lines (TR-544-1 progeny), and 20 plants (untreated cv ‘Tamla’).In conclusion, mutants with altered fatty acid composition could be isolated via EMS-induced mutagenesis. M4 seeds of two mutants (TR-458-2 and TR-544-1) had >75% oleic acid composition with a reduction in linoleic and linolenic acid to an amount equivalent to that of the increase in oleic acid. Selected mutants were superior to the ‘Tamla’ parent, as evident from up to 6% higher oleic acid content. The present study was successful in developing diverse B. napus mutant progenies having high oleic acid content, which is a significant advance towards the development of rapeseed lines for both food and non-food uses. The change in fatty acid profile in the identified progenies may be due to mutations in one or more genes controlling desaturation of oleic acid in these plants. The induced mutants can be utilized as parents in crosses for the development of high-quality rapeseed varieties.
