

서 론
재료 및 방법
실험재료 및 재배방법
과습 처리
생육 특성 조사
통계 분석
단백질 추출
이차원 전기영동
In-gel Digestion
질량 분석 및 bio-informatics을 이용한 단백질 동정
mRNA 발현 분석
결과 및 고찰
팥 유묘기 과습 처리 시 생육특성
과습 처리 시 팥 잎의 단백질 발현
과습 처리 후 팥 잎의 mRNA 발현
결 론
적 요
서 론
팥은 우리나라의 콩 다음으로 재배면적이 큰 두과작물로서 중국 한국 일본에서 전 세계의 생산량의 대부분을 차지하고 있다. 팥은 단백질을 공급하는 주요 식량 자원이며(Sacks, 1977), 주성분은 탄수화물과 단백질이며 무기질, 비타민 등의 영양소와 소량의 사포닌을 함유하고 있다(RDA, 2016). 최근 우리나라는 쌀 생산 과잉과 소비량 감소로 인하여 논토양에서 밭작물을 심는 정책을 실시하고 있다. 콩 자급률 제고 대책의 일환으로 논에서의 콩 재배를 권장하듯 최근에는 팥 또한 기능성 성분이 많이 밝혀짐으로써 소비 증가에 따른 논에서의 팥 재배를 권장하고 있다. 하지만 팥은 습해에 약해 유묘기에 잘록병 발생 등 과습 피해가 크고 이로 인해 수량의 감소가 콩, 수수와 같은 밭작물보다 더 큰 것으로 알려져 있다(Chun et al., 2016). 팥은 파종시기부터 신장기에 장마철이 접어들면서 집중강우로 과습 피해가 우려되며 논에서의 생육 저하를 줄이며 수량 확보를 위한 문제해결이 필요한 실정이다.
Proteomics는 세포나 조직에 있는 모든 단백질의 구조와 기능을 해명하는 연구 분야로, 최근의 proteome이라는 용어는 특정 기관 혹은 조직에서 특정 스트레스에 나타나는 모든 단백질의 발현에도 적용되고 있다(Woo et al., 2003). Proteomics는 단백질 수준에서 유전자 발현에 관한 특성을 알아내고, 외부환경변화에 의한 단백질 발현에 관한 정보를 획득할 수 있는 기술로서 발현된 단백질의 기능 해명에 유용한 기술이라 할 수 있다(Pandey & Mann, 2000).
본 연구는 농촌진흥청 국립식량과학원에서 분리육종법으로 육종된 팥 품종을 이용하여, 팥의 유묘기에 토양 과습에 따른 생육 변화를 알아보고 과습 조건에서 재배한 팥엽의 프로테옴을 대조구와 비교하였다. 또한 단백질 발현 양상을 통해 얻어진 과습 관련 유전자를 선별하여 RNA 발현 수준을 비교 분석하였다. 따라서 팥의 단백질 발현양상 및 RNA 발현양상의 차이를 비교함으로써 팥의 피해와 과습 작용기작에 대한 기초 및 관련 유전자를 발굴하고자 수행하였다.
재료 및 방법
실험재료 및 재배방법
본 실험은 농촌진흥청 국립식량과학원으로부터 분양 받은 중생종 아라리(밀양 8호)를 공시 품종으로 2017년 7월부터 2017년 9월까지 충북대학교 농업생명환경대학 부속 농장 온실에서 수행되었다. 7월 6일에 충북대학교 부속 농장에서 논토양을 2.7 kg 채운 Wagner pot에 6립씩 직파하였으며 발아 10일 후 유묘기에 3본이 되도록 솎아주었다. 논토양 과습 스트레스 실험은 파종 후 18일이 지난 7월 24일에 과습 처리를 시작하여 3일 동안 진행하였으며, 대조구는 관행적 방법으로 재배하였다.
과습 처리
과습 처리는 팥이 심어져 있는 Wagner pot를 큰 통에 넣은 후, pot 밑면의 급수구로부터 수분이 공급되어 지하수위를 표면 5 cm가 되도록 처리하였다. 처리구와 대조구 간의 토양수분량 차이를 실험적으로 확인하기 위해서, 실험구 처리기간 3일 동안 매일 WT-1000N (RF sensor, KOREA)를 이용하여 대조구와 처리구 간의 토양수분량을 3반복으로 측정하여 비교 확인하였다. 유묘기 실험에서 처리구의 토양수분함량은 43∼49%로 유지하였으며, 대조구의 경우 14∼16%로 유지하였다.
생육 특성 조사
생육특성 조사로는 초장, 생체중, SPAD value를 측정하였다. 초장은 과습처리 0일차부터 3일차까지 측정을 하였으며 4일 동안 같은 위치의 초장을 측정하였으며 생체중은 과습처리가 종료되는 날 잎, 줄기, 뿌리 3가지로 나눠 측정하였다. 과습 처리 시작 후 3일간 매일 SPAD-502 plus (Konica Minolta, JAPAN)를 이용하여 유묘기는 1엽의 같은 위치에서 엽록소 양을 측정하였고, 3반복으로 실시하였다.
통계 분석
통계분석은 데이터 결과 값으로부터 기술 통계값을 통계하는 프로그램 중 하나인 R를 이용하여 결과값을 도출했고 데이터는 평균±표준편차로 표시하였다. 모든 측정은 3반복으로 실시하였다. 초장, SPAD value값은 0일차 포함 4일 동안 1일 간격으로 T-test를 수행하였으며 p-value (α= 0.05)값을 바탕으로 과습스트레스에 대한 변화의 유의성을 검정하였다. 생체중의 경우 과습처리 3일차에 T-test를 하였으며 p-value (α= 0.05)값을 바탕으로 유의성을 검정하였다.
단백질 추출
단백질 추출은 파종 후 18일 뒤인 유묘기에 3일간 과습처리한 팥 품종 아라리의 잎을 채취하여 사용하였다. 단백질 추출은 액체질소를 이용하여 효과적으로 파쇄 후 불순물 제거를 위해 TCA 침전법을 이용하였다. 50 ml tube에 10% TCA와 0.07% 2-Mercaptoethanol를 섞어 만 든 용액을 10 ml 채운 후 곱게 갈린 시료를 넣은 후 초음파 처리를 10분간 실시해준 후 원심분리(12000 g, 4°C, 20분)한 후 상등액을 제거하였다. 미리 준비한 2 ml tube에 50 ml tube에 남아있는 하층액과 시료를 잔여물 없이 옮긴다음 아세톤으로 희석한 0.07% 2-Mercaptoethanol 용액을 첨가해준 후 centrifuge (20000 g, 4°C, 5분)를 실시해주고 상등액을 제거하였다. 시료를 세척하기 위해 앞선 과정을 3회 반복 실시해준다. 세척 후 회전감압농축기를 사용하여 건조 시켜준 후 lysis buffer [8M Urea, 2% NP-40, 2% Ampholine (pH 3-10), 5% 2-Mercaptoethanol, 5% Polyvinylpyrrolidone (PVP-40)] 를 추가하여 1시간 동안 상온에서 voltexing하여 반응시켜 주었다. 그 후 원심분리(20000 g, 25°C, 20분)하여 상층액을 새로운 1.5 ml 튜브에 옮겨주었다. 모든 실험은 3반복으로 실행되었으며 추출된 단백질 시료는 Bradford (1976)법으로 분석을 하여 단백질 농도를 결정하였다.
이차원 전기영동
일차원 전기영동(one-dimentional electrophoresis: 1-DE)은 13 cm 유리관에 straw gel [Urea, 30% acrylamide, 10% NP-40, Ampholine (pH 3-10), 10% APS, TEMED]을 넣어 6시간 이상 굳힌 후 추출한 단백질 400 μg을 기준으로 하여 straw gel 위에 주입하여 전기영동 하였다. 전압은 150V에서 1시간, 300V에서 1시간, 500V에서 16시간으로 전압을 증가 조정하여 전기영동 하였다. 일차원 전기영동 후 straw gel을 buffer solution (시료용 완충액) [glycerol, 0.5 M Tris (pH 6-8), 10% SDS, 2-Mercaptoethanol]에 담가 15분씩 4회 buffer solution을 교체해주며 gel을 세척하며 평형화 해주었다. 2차 전기영동 분석을 위해서 12% separating gel [D.W, 1.5M Tris (pH 6-8), 30% acrylamide, 10% APS, 10% SDS, TEMED], 5% Staking gel [D.W, 0.5 M Tris (pH 6-8), 30% acrylamide, 10% APS, 10% SDS, TEMED] 에 전기영동을 실시하였으며 전류는 시료당 35 mA로 걸어주었다. 2차원 전기영동이 완료되면 separating gel을 coomasie brilliant blue (CBB G-250)로 염색해주었고 염색 후 각각의 gel은 scanner (HP Scanjet G4010, USA)을 이용하여 스캔했다. 각각의 2D-gel 이미지 분석은 Progenesis samespot program (Nonlinear Dynamics, Newcastle-upon-Tyne, UK)을 사용하여 분석, 실행하였다.
In-gel Digestion
이미지 분석의 결과를 바탕으로 팥 2엽기(유묘기) gel에서 protein spot들을 선정하였다. 각각의 gel에서 선정된 protein spot들을 2-D gel에서 1 × 1 mm 크기로 절단한 후 1.5 ml tube에 옮겨준다. 30% MeOH을 사용하여 gel조각을 세척한 후 destaining buffer [50% ACN/10 mM ammonium bicarbonate]에 20분간 shaking을 하였으며 Dye 색깔이 gel에서 완전히 사라질 때까지 앞선 과정을 2-3번 실시하였다. Gel을 건조한 후 tube에 reduction buffer (100 mM ABC에 10 mM DTT를 넣은 용액)에 56°C에서 1시간 반응시켜준 다음 alkylation buffer [100 mM ABC에 55 mM IAA 넣은 용액] 에 40분간 반응시켜 주었다. Alkylation buffer를 완전히 제거해주고 각각의 tube에 100% ACN으로 세척 후 air-dry 해준다. Tube에 Trypsin buffer [trypsin + 80 µl resolving buffer (acidic)]를 처리하여 30분간 4°C 냉장보관 후 37°C에서 16시간 incubating 한 후에 상등액을 옮겨 담고 50% ACN/0.1% FA 50 µl를 첨가 후 상등액을 취한 다음 speed vacuum centrifuge을 이용하여 완전히 건조 시켜 최종 질량 분석을 위한 시료로 사용하였다.
질량 분석 및 bio-informatics을 이용한 단백질 동정
Peptide 확인을 위해 Finningan LTQ-FT-ICR mass spectrometer (Therm Electon, Bremen, Germany)로 구성된 Nano-LC/MS 시스템에서 단백질 질량 분석을 실시하였다. 각 시료들을 탈염하고 시료 10 µl를 auto sampler를 사용하여 C18 trap-column (I.D., 300 µl, length 5 mm, particle size 5 µl, LC Packings)에 주입하여 20 µl/min의 flow rate로 농축하였다. 이후 MS와 MS/MS acquisition을 반복하며 data-dependent mode로 작동하였고, 실험 조건(Condition: 2.2 kV spray voltage, 220°C ion transfer tube temperature, 1.3 millitors collision gas pressure)올 설정하였다. Ion 검출은 500 counts를 한계점으로, 활성화는 q = 0.25, 활성화 시간은 30 ms로 적용하였다. 동정된 단백질 peptide의 MS spectra data값은 먼저 MASCOT ver 2.0 (Matrix Science, UK)의 database에서 검색하였으며, peptide 오차범위는 50 ppm, fragment ion 오차 범위는 0.5 DA로 설정하였다. 검색 결과를 토대로 수집한 protein dascription과 peptide sequence 정보를 PIR (http://pir.georgetown.edu)에서 protein sequence를 검색하였다. 최종 확인된 팥 잎의 단백질은 분자량과 pI 2가지 항목을 통해 각각의 특성에 따라 분류하여 정리하였다.
mRNA 발현 분석
RNA 추출 및 cDNA 합성
RNeasy Plant Mini Kit (QIAGEN) 를 사용하여 total RNA을 추출하였으며 추출한 RNA의 농도를 측정하기 위해 Nano Drop One (Thermo fisher scientific, USA)을 사용하였고, cDNA 합성을 위해 ReverTra Ace® qPCR RT Kit/Master Mix (Toyobo, Japan)를 사용하였다. 추출된 RNA의 최종 농도는 Nuclease-free water를 첨가하여 1 ng/µl로 희석시킨 후 65°C에서 5분간 처리하였다. RNA를 담은 1.5 ml 튜브를 얼음에서 식혀준 후, DNaseⅠ과 Oligo dT, Random primer가 함유된 4× master mix 2 µl과 Nuclease-free water, RNA 6 µl을 첨가하여 최종 부피를 8 µl로 조절하여 37°C에서 5분간 처리하였다. PCR 증폭은 BioRad T100 Thermal Cyler (Bio-Rad, USA)를 사용하였고, PCR은 37°C에서 15분, 50°C에서 5분, 98°C에서 5분으로 설정하여 Reverse transcription을 진행하였다.
실시간 중합효소 연쇄 반응(Real-Time PCR) 분석
RT-PCR을 통하여 target 유전자가 증폭되는 것을 확인한 후, target 유전자의 mRNA 발현량 분석을 위해 qRT-PCR을 수행하였다. 합성한 cDNA 2 µl와 SYBR® Green KOD gPCR 4.8 µl, 50x ROX Dye 0.2 µl, 각각의 프라이머 1.5 µl를 혼합한 뒤 멸균된 증류수로 최종 부피를 총 10 µl로 조절하여 사용하였다. PCR 반응은 50°C로 10분간 진행 한 후 95°C에서 5분간 predenaturation을 시행한 후 95°C에서 10초간 denaturation, 55°C에서 30초간 annealing, 55°C에서 30초간 extension하고, scanning하는 과정을 60회 반복하였으며, melting curve 분석은 65∼95°C에서 0.5°C씩 5초간 실행하였다.
결과 및 고찰
팥 유묘기 과습 처리 시 생육특성
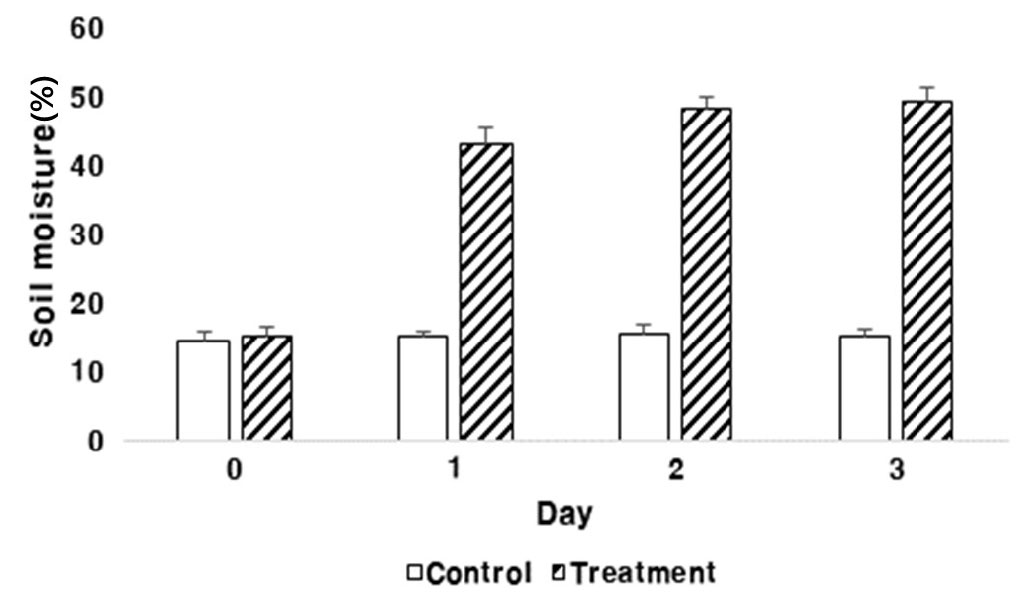
팥 유묘기 토양의 과습 처리 3일 동안 토양수분을 측정한 결과 유묘기 0일차는 대조구 14.5%, 처리구 15.08%, 1일차는 대조구 15.08%, 처리구 43%였다. 2일차는 대조구 15.48%, 처리구 48.45%, 3일차는 대조구 15.11%, 처리구 49.48%의 토양수분이 측정되었다(Fig. 1). 따라서 대조구의 토양수분은 10-20% 사이었으며 처리구는 40-50% 사이로 과습 조건으로 확인되었다.
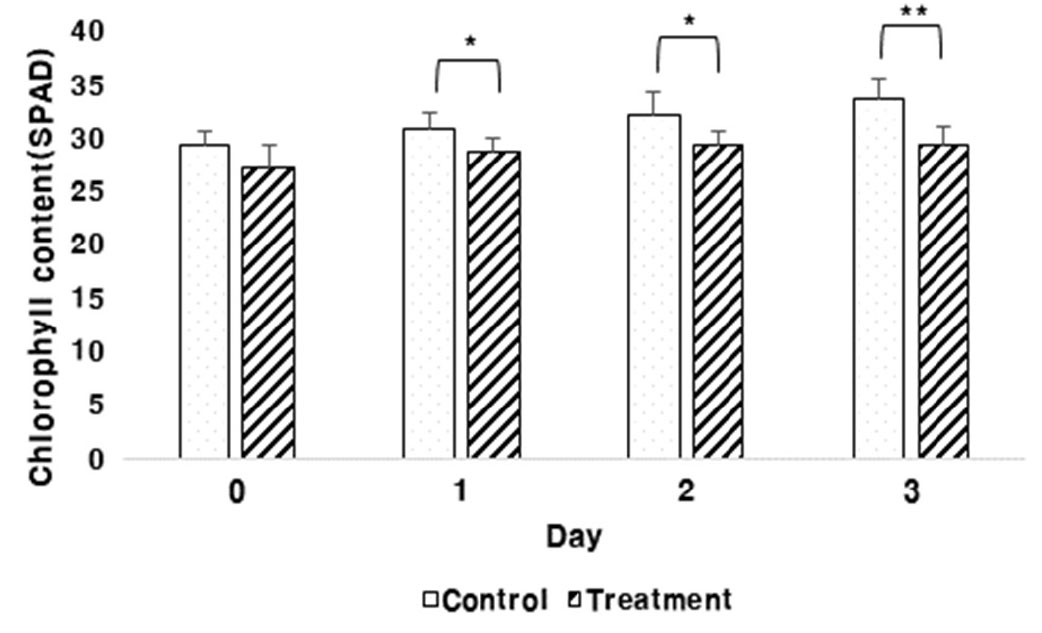
팥 유묘기 토양의 과습 처리 3일 동안 측정한 SPAD value 값을 측정한 결과는 Fig. 2와 같다. 과습 처리 3일 동안 처리구는 SPAD value의 증가율이 낮았던 반면 대조구는 처리구에 비해 증가율이 높았다. 과습처리 1일과 2일차에서 SPAD value가 통계적으로 유의하다는 결과가 나왔으며 특히 3일차에 대조구와 처리구간의 SPAD value 값의 차이가 커 고도로 유의한 결과를 나타냈다. 따라서 과습으로 인해 유묘기인 팥이 생리적으로 스트레스를 받은 것으로 판단되었다. 이는 침수처리 시 고추 속 작물의 광합성률은 3일째 50% 이하로 감소하였다는 보고(Ou et al., 2011)와 같이 토양의 과습 상태가 지속됨에 따라 작물에 대한 과습 스트레스가 작물의 지상부까지 영향을 주어 엽록소 피해로 이어지는 것으로 사료되었다.
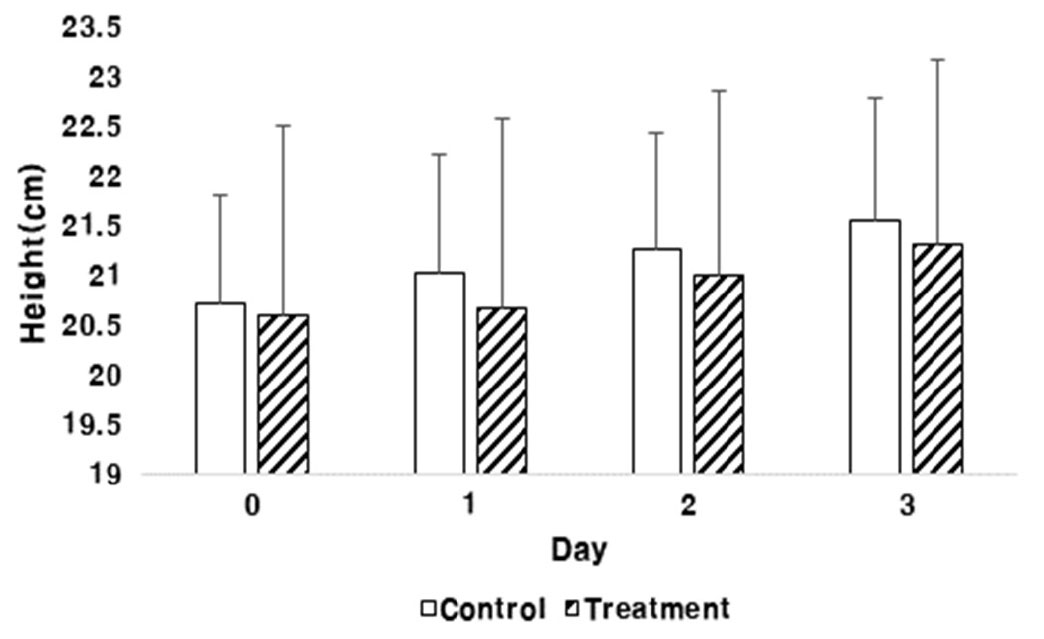
초장은 과습 처리 1일차부터 대조구에 비해 처리구에서 생장이 지연되는 경향을 보였으나 과습 처리 날짜 별 T-test 결과 통계적으로 유의성은 보이지 않았다(Fig. 3). 토양의 배수조건이 매우불량인 논과 약간불량이 논에서의 조, 수수, 기장의 생육특성을 조사한 결과 과습에 의한 생육장해는 간장에 영향을 크게 미치는 것으로 나타났다(Jung et al., 2013).
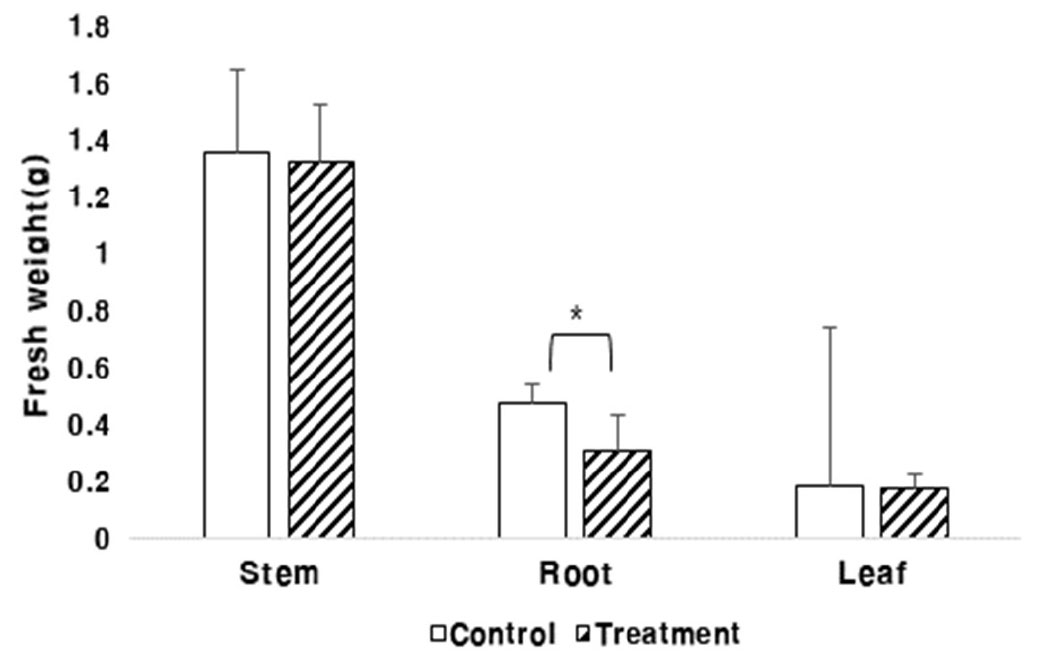
생체중(줄기, 뿌리, 잎)에서도 초장과 비슷한 결과로 과습처리가 끝난 후 측정한 결과 대조구에 비해서 처리구의 생장이 저조하였고, 뿌리의 경우 과습 처리시 생장이 대조구보다 확연히 떨어지는 것을 볼 수 있었다(Fig. 4). 이는 과습으로 인해 식물체의 양분흡수와 물질이동이 감소한(Bacanamwo & Purcell, 1999) 결과로 판단되었다. 뿌리 생체중은 과습에 의한 차이를 보였으나, 줄기와 잎은 과습의 영향을 확인할 수 없었다. 따라서 팥의 유묘기에 과습으로 인한 피해를 받으면 생육이 저조하고 이는 수량에 영향을 미치므로 팥의 생육이 가장 왕성한 장마기간에 피해를 최소화하기 위해 이랑을 높이는 등 과습 피해를 예방하는 것이 중요할 것으로 판단되었다.
과습 처리 시 팥 잎의 단백질 발현
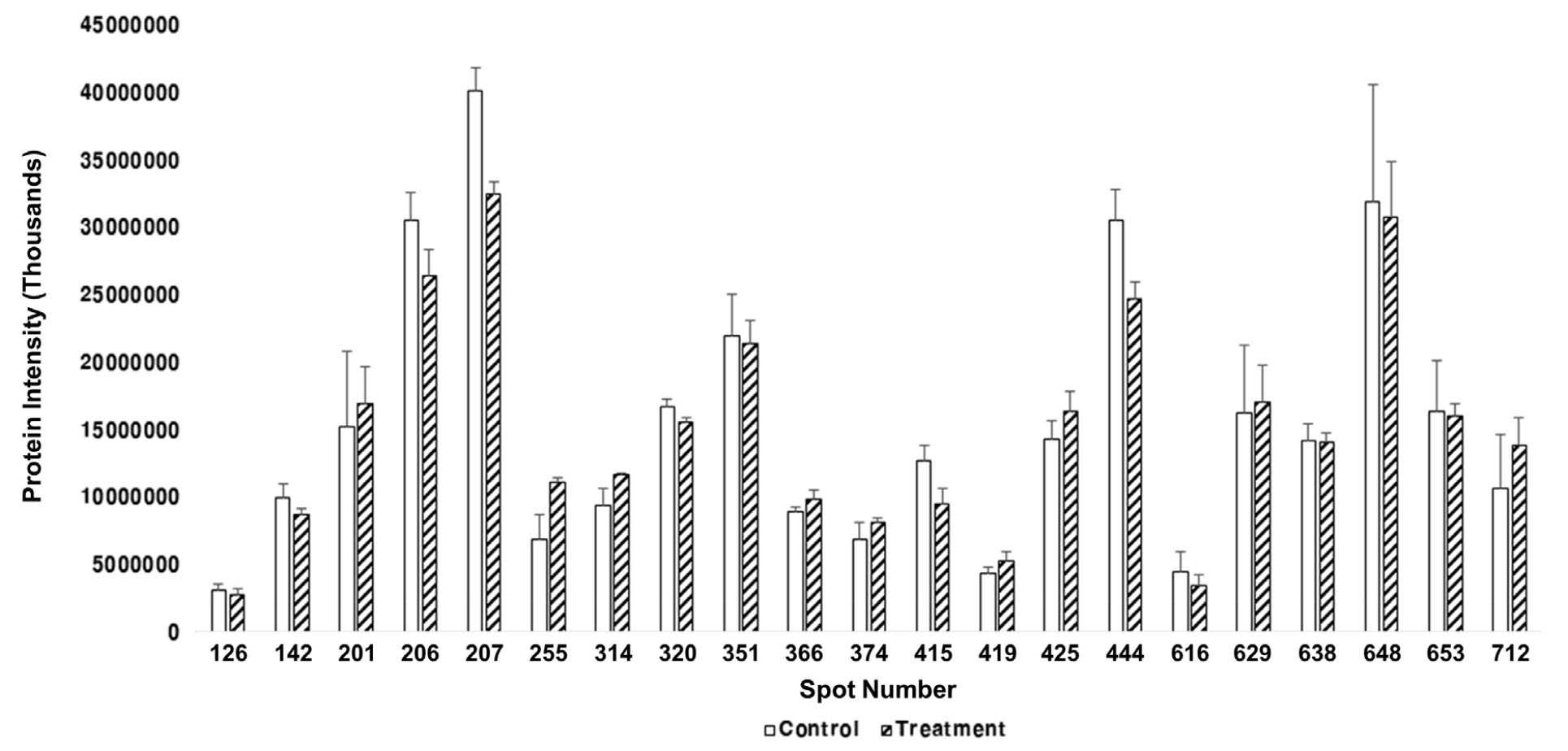
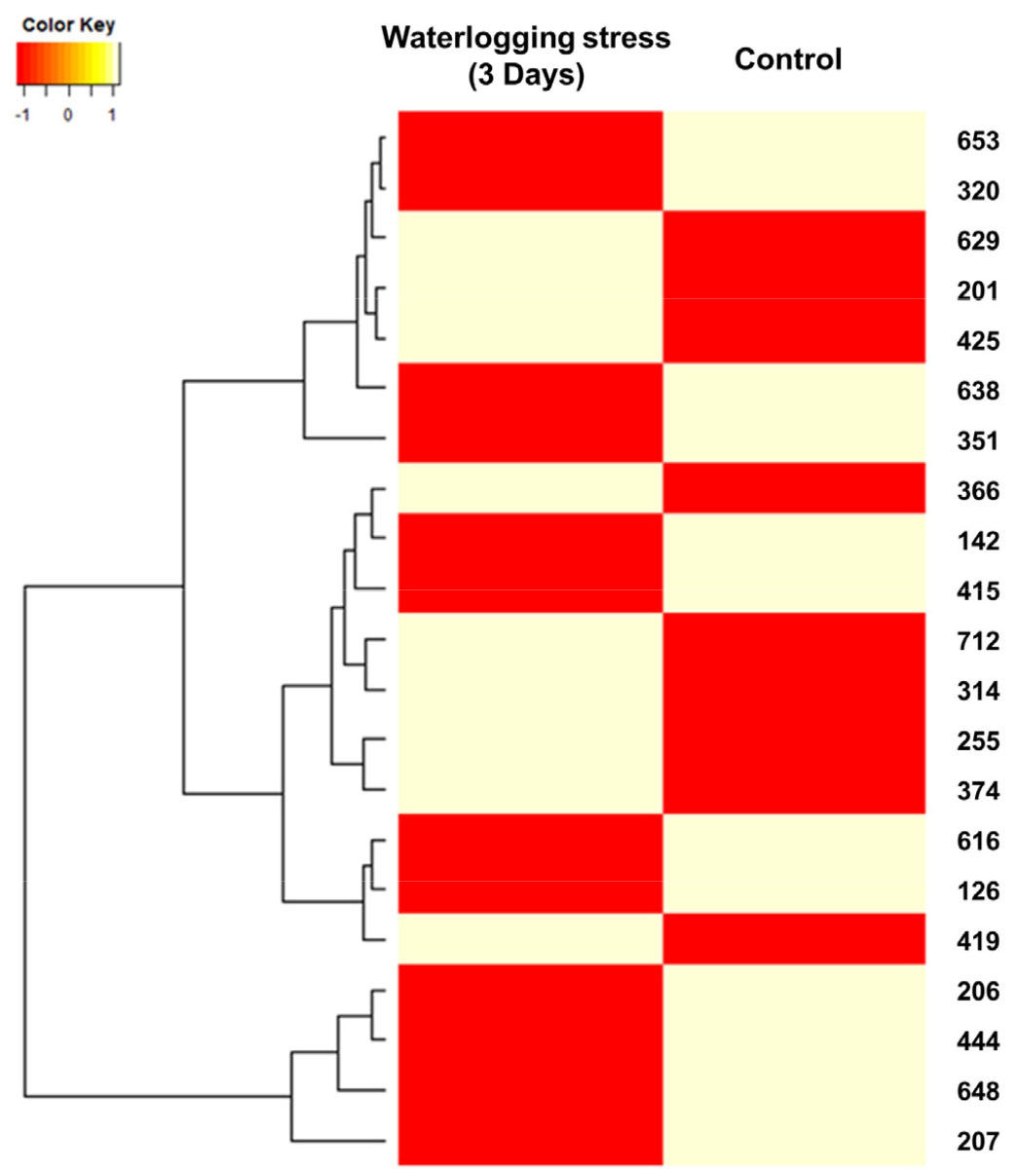
과습 처리 3일 후 잎에서 추출한 단백질 발현 양상을 확인하기 위하여 Progenesis samespot software program을 사용하여 단백질 spot을 분석한 결과 2-D Gel에서는 366개의 단백질 spot이 검출되었고 그 중 차별적으로 발현된 21개의 단백질을 선택하였다. 이미지 분석 시 기준이 된 Reference gel을 Mass spectrometry로 분석하였고, 각각의 spot은 구획을 원으로 나누었고 numbering 하여 선택하였다(Fig. 5). 단백질 중 9개가 up-regulated 되었고 12개가 down-regulated 되었다(Fig. 6). 유묘기 과습 처리한 대조구와 처리구간에 상향 발현된 단백질과 하향 발현된 단백질간의 관계를 히트맵으로 시각화하였다(Fig. 7).

Fig. 5.
Reference 2-DE gel map of cultured adzuki bean leaves after 3 days of waterlogging stress. Proteins were extracted from adzuki bean leaves, separated by two-dimensional gel electrophoresis (2-DE), and visualized by Coomassie brilliant blue staining. (C) Control, (T) Waterlogging for 3 days. The red arrows (↑) denote proteins that increased in abundance and the blue arrows (↓) denote proteins that decreased in abundance.
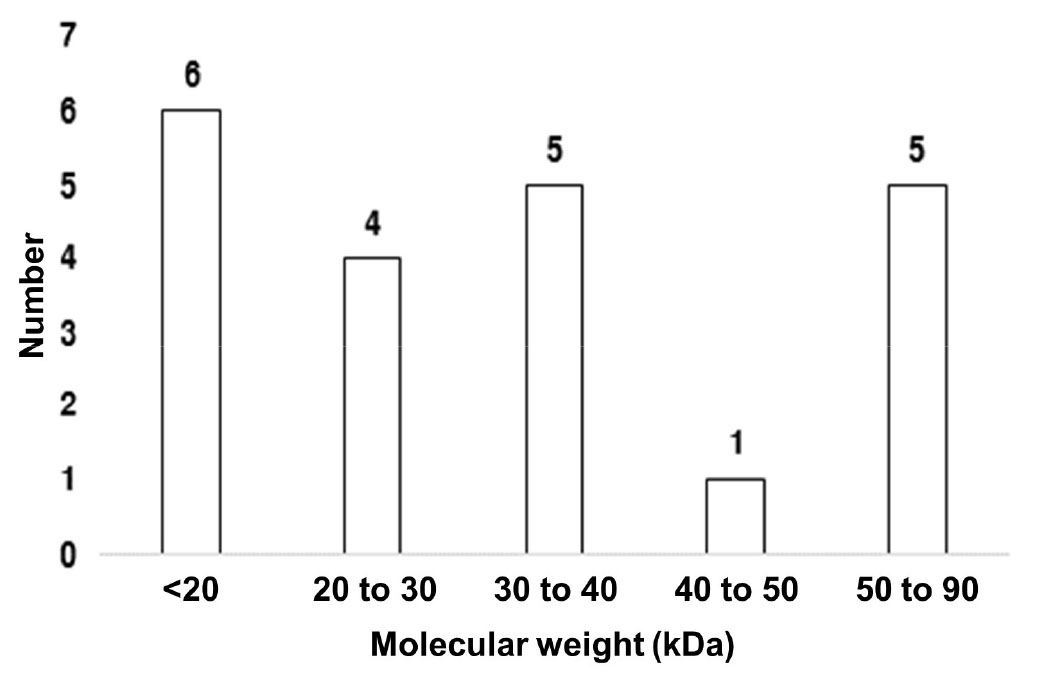
단백질을 분자 크기로 비교한 결과 21개의 단백질의 분자량은 10 kDa∼90 kDa에 속하는 것으로 확인되었다. 그 중 10 kDa∼50 kDa에 속하는 spot은 76%이고 50 kDa∼90 kDa에 속하는 spot이 24%였다(Fig. 8).
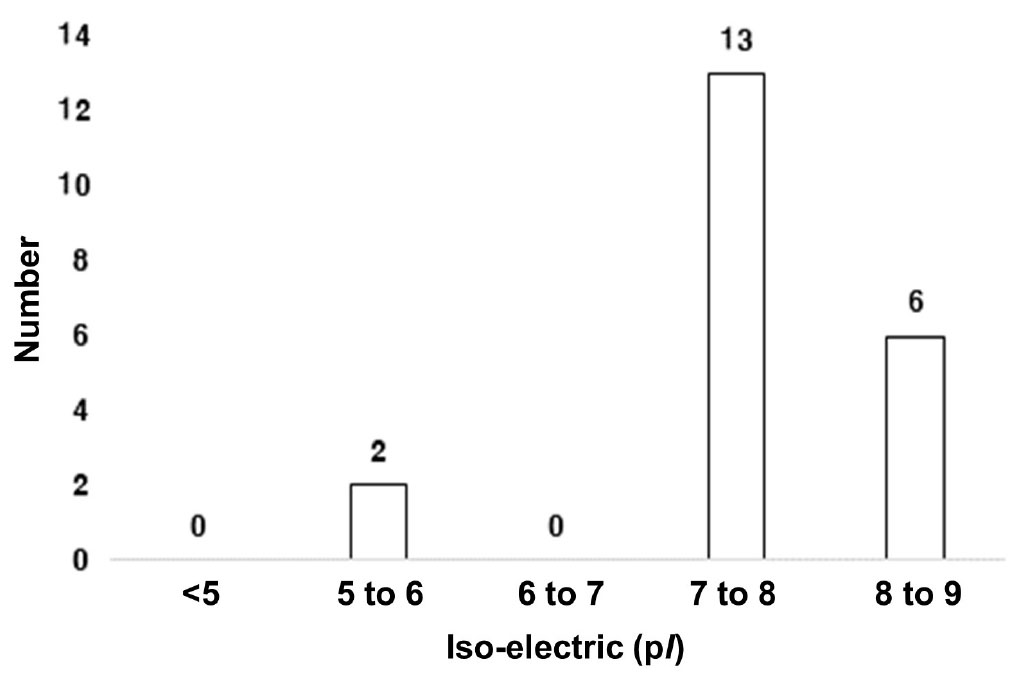
등전점(pI)의 경우, 가장 높은 단백질 분포는 pI가 7∼8 사이에서 62%였고 8∼9 사이는 28%로 중성인 단백질이 유묘기에서는 가장 많이 나왔으며 염기성 단백질이 많이 분포되어 있음을 알 수 있었다(Fig. 9).
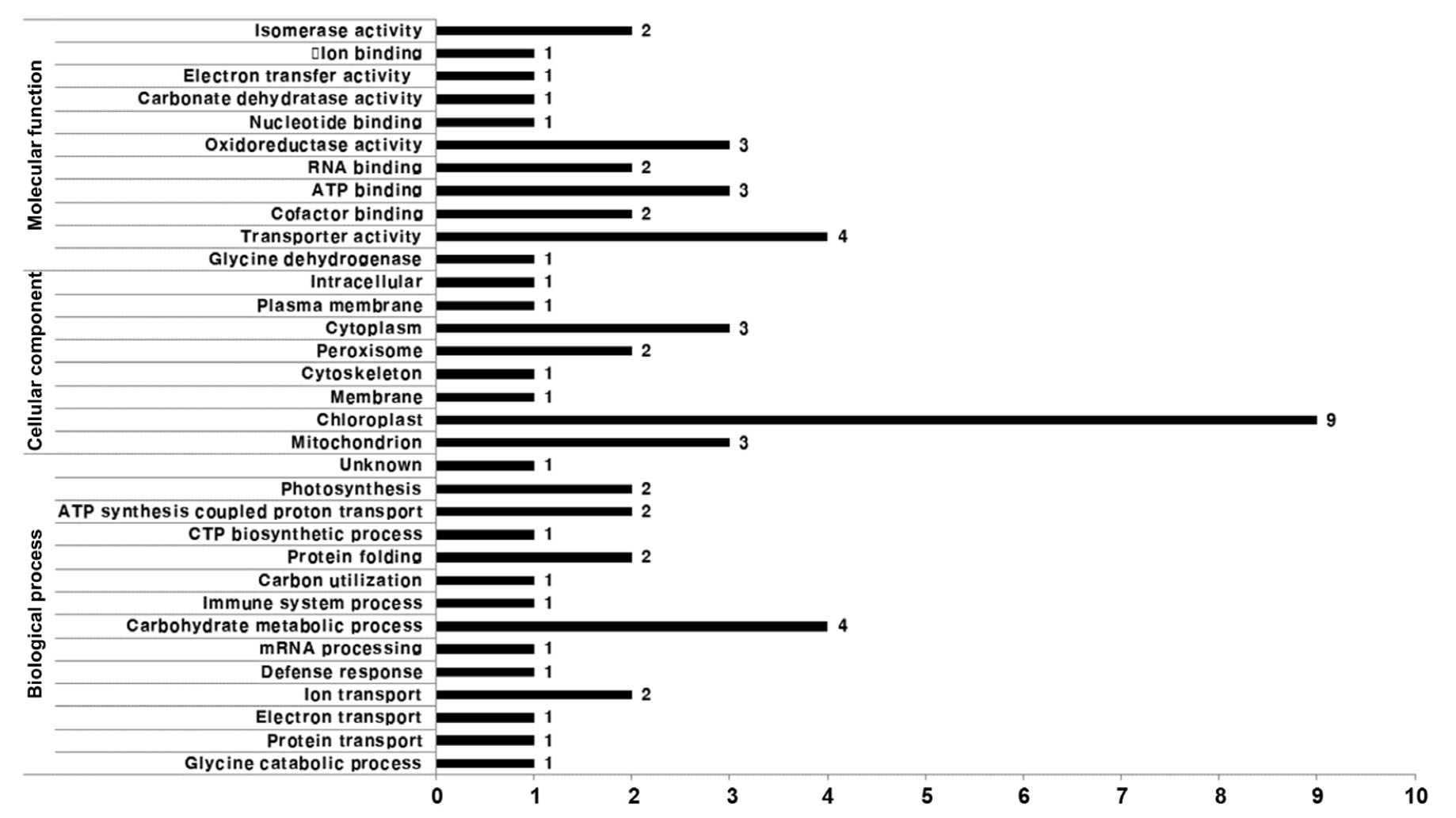
LTQ-FT-ICR MS를 사용하여 차별 발현된 21개의 단백질을 동정하였으며, 확인된 단백질들은 Table 1과 같다. 확인된 21개의 단백질을 molecular function, cellular component, biological process를 기준으로 분류하였다(Fig. 10). Molecular function 에서는 Transporter activity가 19%로 가장 많았으며 그 다음으로는 ATP binding, Oxidoreductase activity가 14%로 나타났다. Cellular component에서는 Chloroplast가 43%로 가장 많았고 그다음으로 Mitochondrion, Cytoplasm이 14%로 나타났다. Biological process에서는 Carbohydrate metabolic process가 19%로 가장 많았는데 이 과정에서 나온 단백질은 모두 과습 스트레스와 관련 있는 단백질이었다. 이것으로 보아 탄수화물 대사 과정에 적용되는 단백질들이 팥 유묘기에 과습스트레스로 인해서 많은 영향을 받는 것으로 판단된다.
Table 1.
List of differentially expressed proteins identified in the leaves of adzuki beans at the two-leaf stage under waterlogging stress using LTQ–FT–ICR MS.
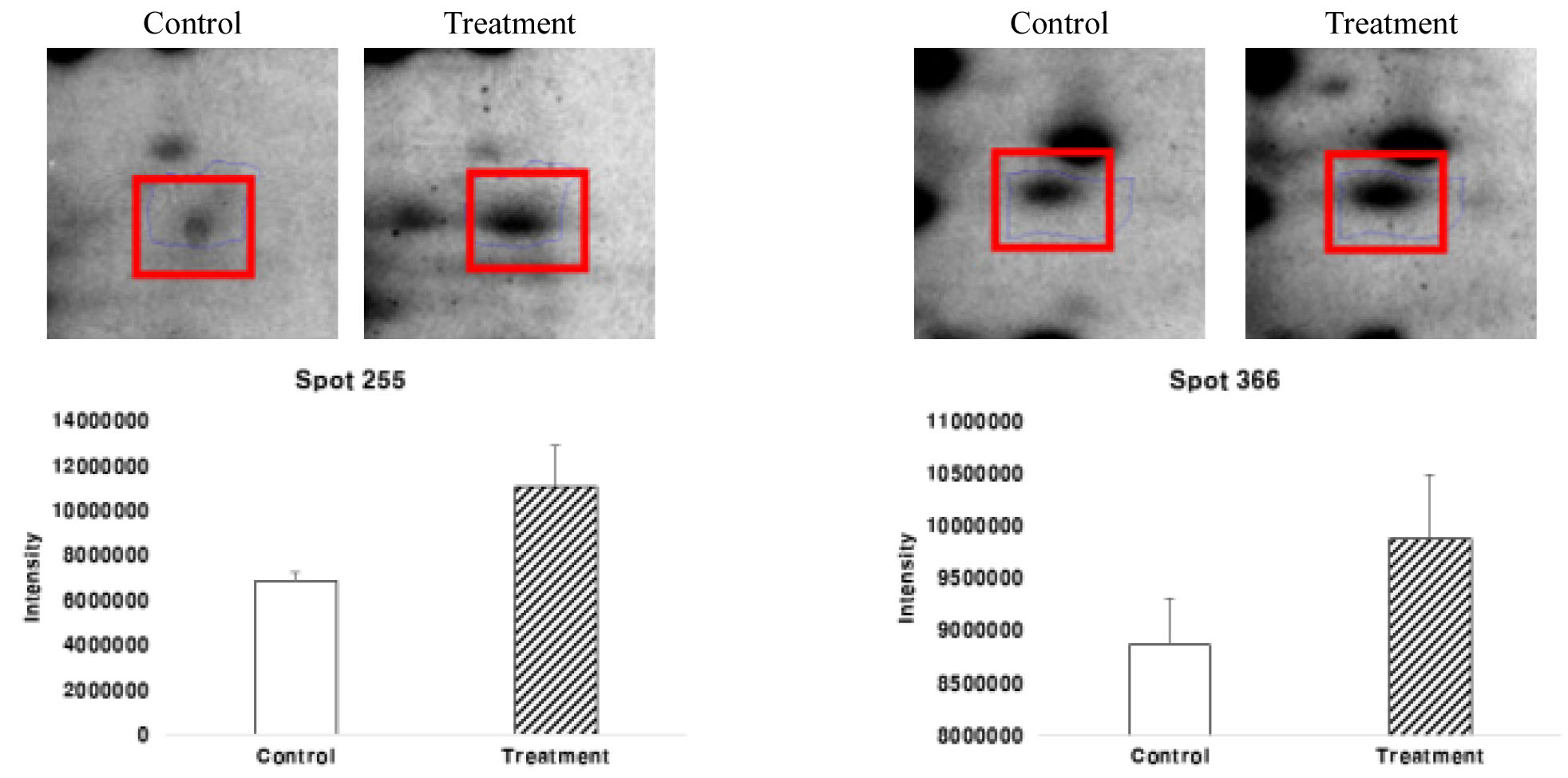
Spot number 255 Maturase K (matK)는 regulatory protein으로 전사 과정에서 RNA 중합효소에 의해 전사되는 DNA 분자 영역에 영향을 미치는 단백질이다. 이러한 단백질은 세포에서 단백질 합성을 조절하는데 도움을 준다. matK는 5’와 3’ 사이에 중첩된 엽록체 코딩 유전자이고 엽록체 게놈의 큰 단일 copy 영역에서 trnK, tRNA-lysine의 엑손이다(Sugita et al., 1985). matK 진화의 방식과 속도는 다른 엽록체 유전자와 구별된다. matK에서의 뉴클레오타이드 치환률은 Rubisco의 큰 하위 단위(rbcL)보다 3배 높고 아미노산 치환률은 6배 더 높다(Johnson & Soltis, 1994; Olmstead & Palmer, 1994). 이 높은 뉴클레오타이드 및 아미노산 치환률은 모든 분류학적 수준에서 식물 간의 진화 관계를 해결하기 위한 신호를 제공한다. 높은 치환률에 덧붙여, matK는 다양한 숫자와 크기의 indel을 보여주고 대부분의 indels은 reading frame을 보존하면서 3의 배수로 나타내었다(Hilu & Liang, 1997; Soltis & Soltis, 1998; Hilu et al., 2003). 그러나 일부 식물 군에서 발견되는 indels의 존재, 높은 치환률 및 조기 정지 코돈은(Kores et al., 2000; Kugita et al., 2003) 이러한 특징을 지닌 유전자가 단백질 구조와 기능을 안정하게 유지해준다. matK는 추정 maturase 도메인을 포함하는 고등 식물의 엽록체 게놈에서 발견되는 유일한 유전자이며(Neuhaus & Link, 1987), 이 matK의 단백질 발현 수준은 처리구에서 대조구보다 단백질 발현양이 증가한 것을 확인 할 수 있었다(Fig. 11). 이러한 이유는 과습 스트레스에 따라 저항할 수 있는 단백질을 세포에서 합성 조절하는데 이용되어야 하기 때문에 처리구에서 증가한 것이라고 사료된다.
식물 신진대사와 관련된 Spot number 366 Malate dehydrogenase 2 (MDH)는 MDH gene을 가지는 단백질이다. MDH는 과습 스트레스에 대한 식물 반응의 지표로 사용되어왔다(Daugherty & Musgrave, 1994). 말산 탈수소 효소는 L-말산(Malate)으로부터 수소를 제거하고 옥살로아세트산(Oxaloacetate)과 NADH를 생성하는 반응에 촉매하는 효소이자 TCA 회로 대사계에 관여하는 효소로 중요하다. Malate dehydrogenase는 총 포도당 산화에서 마지막 NADH2의 형성을 촉진한다(Rocha et al., 2010). Malate dehydrogenase 2 (MDH)는 대조구보다 처리구에서 더 많이 발현이 되었다(Fig. 11). 이것으로 보아 MDH는 과습 스트레스에 노출이 되었을 때 단백질의 피해를 줄이기 위해 antistress 기능을 하는 단백질이라 사료된다.
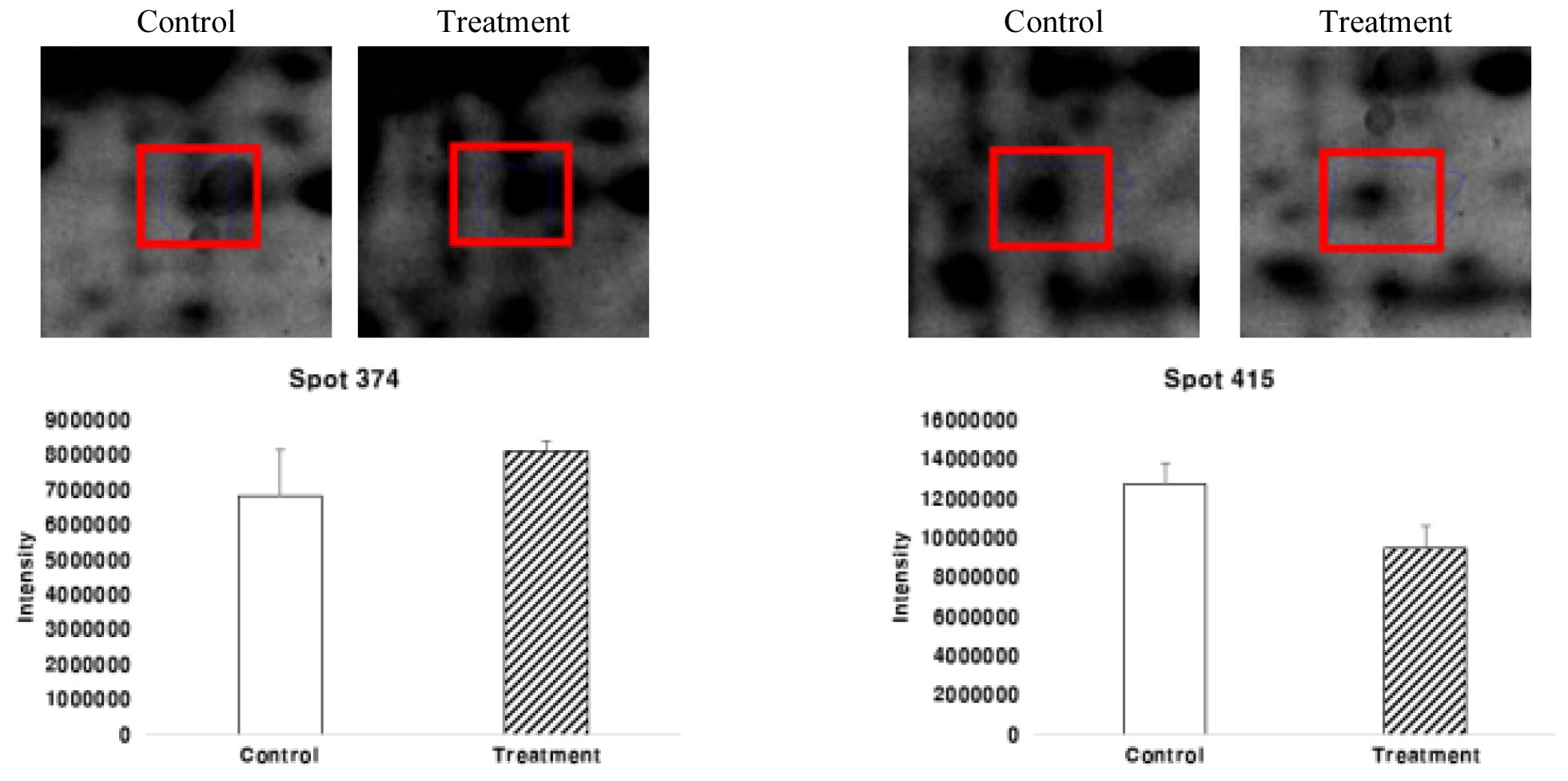
Spot number 374 Glyceraldehyde-3-phosphate dehydrogenase (GAPDH)는 본 연구에서 확인이 되었는데, GAPDH는 당분 분해 과정의 첫 번째 단계에서 D-glyceraldehyde 3-phosphate를 3-phospho-D-glyceroyl phosphate으로 전환시키는 주요 효소로 세포 내 ATP 수준과 탄수화물 신진 대사의 유지를 위한 필수 핵심 효소이다. GAPDH 단백질은 대조구에 비해 처리구에서 발현량이 증가하였다(Fig. 12). 이것은 과습 스트레스가 세포 내 탄수화물 대사에 영향을 미치는 것이라 확인이 되었다.
Spot number 415 Carbonic anhydrase (CA)는 광합성 관련 단백질로써 CO2와 HCO3의 상호전환을 촉진하는 효소이다(Khalifah, 1971). CA는 광합성과 호흡 모두를 포함하여 카르복실화 또는 탈 카르복실화 반응을 포함하는 많은 생리학적 기능에 중요하다. 또한 CA는 활발하게 광합성 세포에 무기 탄소 운반에 참여하거나 능동적으로 호흡하는 세포부터는 떨어진다는 점은 분명하다(Henry, 1996).
CA 단백질은 처리구보다 대조구에서 많이 발현이 되었는데(Fig. 12) 세포 표층에서 수중의 HCO3-를 외견상 취입하는데 작용하고, 엽록체 내의 것은 카르복시 불균등화 효소로의 CO2공급을 촉진하는 작용을 하는 것으로 생각되었다. 따라서 과습스트레스에 따라 CO2의 공급을 줄이기 위해 처리구에서 CA의 발현양이 줄어든 것으로 사료된다.
과습 처리 후 팥 잎의 mRNA 발현
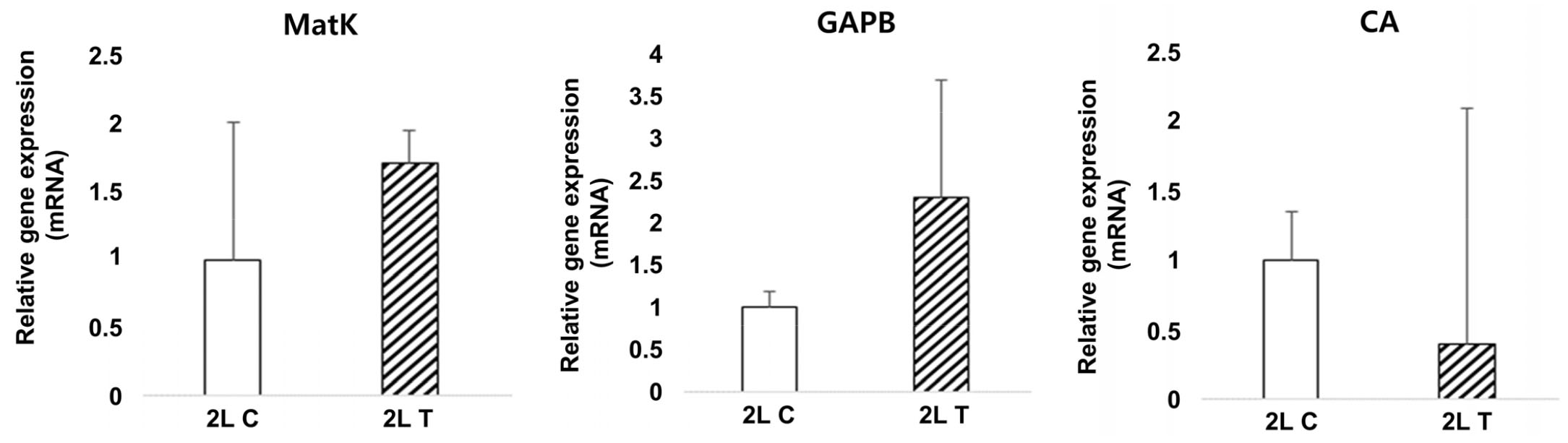
유묘기의 과습스트레스에 따른 MatK (spot number 255) 유전자의 mRNA는 대조구보다 처리구에서 발현양이 증가하여 단백질 결과와 유사한 경향을 보였다(Fig. 13). MatK는 regulatory protein으로 전사 과정에서 RNA 중합효소에 의해 전사되는 DNA 분자 영역에 영향을 미치는 단백질로 세포에서 단백질 합성을 조절하는데 도움을 준다.
GAPB (spot number 314) 유전자의 mRNA 발현양을 확인한 결과 단백질 발현과 비슷하게 처리구에서 발현양이 증가한 것으로 나타났다(Fig. 13). GAPB는 당분 분해 과정의 첫 번째 단계에서 D-glyceraldehyde 3-phosphate를 3-phospho-D-glyceroyl phosphate으로 전환시키는 주요 효소로 과습스트레스가 세포 내 탄수화물 대사에 영향을 미치는 것으로 확인이 되었다.
CA (spot number 415) 유전자는 단백질 발현에서 과습에 의해 감소하였는데 mRNA 발현양도 처리구에서 감소한 결과를 보였다(Fig. 13). CA는 탄산탈수효소라고도 하며 기능은 탄소고정과정에서 루비스코(Rubisco) 효소의 카복실화 반응(carboxylation)을 증가시키기 위해서 엽록체 내에서 이산화탄소 농도를 높이는 것이다. 따라서 광합성과 관련된 유전자들, 특히 엽록소 합성에 관여하는 유전자들이 24시간 동안 과습 스트레스 처리된 목화 식물체의 잎에서 하향 조절되었다는 보고(Christianson et al., 2010)와 유사한 결과로 본 연구에서도 감소하였다.
결 론
팥 유묘기 3일간 과습 처리 시 잎의 단백질 발현양상을 확인한 결과 대조구와 처리구간의 특이하게 발현된 단백질이 21개 확인되었다. 그 중 Maturase K, Glyceraldehyde-3-phosphate dehydrogenase 단백질은 모두 과습스트레스와 연관이 있는 단백질이었고 과습스트레스 조건하에서 증가한 것을 확인할 수 있었다. 이외에도 Malate dehydrogenase 1이 증가되었는데 이 MDH는 과습스트레스에 대한 식물반응의 지표로 사용되어 왔고, TCA회로에 관여하는 효소로 과습 스트레스를 받았을 때 식물체 자신을 보호하기 위해 antistress 기능을 한 것이라 사료되었다. 광합성과 관련된 carbonic anhydrase (CA)는 대조구에 비해 과습 처리구에서 감소발현 되었는데 CA는 광합성 세포에 무기탄소를 운반하는 역할을 하거나 능동적으로 호흡하는 세포로부터 멀어지는 기능을 가지고 있으며 CO2 공급을 촉진하는 작용을 한다. 따라서 과습 처리구에서의 CA 발현 감소는 과습으로 인한 호흡의 문제로 CO2의 공급의 문제로 광합성에 영향을 미치는 것으로 사료되었다.
적 요
논 토양 조건에서 팥의 생육기 중 초기 생육 단계에 해당되는 유묘기에 과습 스트레스 처리 후 생육특성 변화 및 팥 잎의 단백질 발현 양상을 조사한 결과는 다음과 같다.
1. 아라리(밀양 8호)를 공시 품종으로 과습 스트레스 실험을 진행하였다. 3일간 과습 처리하는 동안 팥의 초장과 생체중(잎, 줄기)은 통계적으로 유의하지는 않았으나 생체중(뿌리)와 SPAD value 값은 통계적으로 유의한 결과가 나왔다.
2. 과습 처리 3일 후 잎의 단백질 발현 양상을 확인한 결과 21개의 차별 발현된 단백질 중 과습 스트레스 처리에 의해 9개의 단백질들이 up-regulated 되었고, 12개의 단백질들이 down-regulated 되었다. 단백질을 기능별로 분류한 결과 Biological Process에서 carbohydrate metabolic process와 관련된 단백질들이 가장 많이 나왔고 Cellular Component에서는 Chloroplast에서 가장 많이 존재하였다.
3. Regulatory protein과 관련된 Maturase K 단백질은 처리구에서 대조구보다 발현양이 증가한 것으로 나타났다.
4. Carbohydrate metabolic process와 관련된 Malate dehydrogenase 단백질과 Glyceraldehyde-3-phosphate dehydrogenase 단백질은 과습 스트레스를 받았을 경우 단백질 발현양이 증가하였다.
5. Photosynthesis와 관련된 Carbonic anhydrase (CA)는 처리구보다 대조구에서 단백질 발현량이 더 증가한 것을 확인할 수 있었다. 스트레스와 관련된 단백질 Superoxide dismutase는 처리구에서 대조구보다 단백질 발현양이 증가한 것을 확인할 수 있었다.
