

INTRODUCTION
MATERIALS AND METHODS
Measurement of seed characteristics
Observation of mitotic chromosomes
Observation of stomatal traits and leaf structure
PCR with EST markers
RESULT AND DISCUSSION
Seed characteristics of E. humidus
Leaf structure and stomatal traits of E. humidus compared to common wheat
Identification of chromosomes and polymorphism of E. humidus versus common wheat
INTRODUCTION
Common wheat is a major crop grown worldwide. Recently, wheat consumption in Korea has increased continuously (RDA, 2020). Therefore, wheat is perceived as the second staple food, although Korean wheat has low self-sufficiency (MAFRA, 2021). In this context, improving wheat self-sufficiency is critical for food security in Korea. However, due to climate change, abiotic stresses such as drought and flooding negatively affect the growth and development of wheat, leading to a decrease in wheat yield potential, even when the wheat cultivation area is finite because of double cropping for rice transplantation (Zhang et al., 2022).
One of the methods to improve wheat quality against abiotic stresses is using wild plants as genetic resources because they harbor environmental adaptability; therefore, the use of wild plants is being studied (Zhang et al., 2017; Li et al., 2019; Li et al., 2021). Among many wild plants, Elymus L. is distributed in many countries, according to the Royal Botanic Gardens, Kew. In addition, the genus, comprising approximately 150 species, is the largest among Triticeae plants (Dewey, 1984; Sasanuma et al., 2002). Elymus species have been used to study drought, salt tolerance, cold stress, and biotic stresses (Sasanuma et al., 2002; Zabala et al., 2011; Gong et al., 2019; Khan et al., 2022). Gong et al. (2019) reported that wheat-Elymus translocation lines harbored high levels of Fusarium head blight resistance. Similar to these studies, wild plants are useful genetic resources for improving wheat quality.
Elymus humidus is a perennial plant distributed by its rhizome, while common wheat is an annual plant. In addition, E. humidus, an indigenous plant to Japan, is a useful genetic resource for humidity resistance due to its high adaptability to paddy fields (Sasanuma et al., 2002). Recently, it was reported that E. humidus grows spontaneously around riversides in Korea by Choi et al. (2021). Water stress, such as low moisture or waterlogging, negatively affects and damages wheat growth, causing a decrease in yield potential (Akram, 2011; Ploschuk et al., 2018; Zhao et al., 2020; Komatsu et al., 2022). Although E. humidus exhibits environmental adaptability in wetlands, studies on this species are purely based on limited growth and distribution areas.
In this study, we investigated stomatal traits and leaf structure as cytological characteristics of E. humidus because waterlogging is related to leaf area, chlorophyll content, and stomatal conductance and morphology (Rodríguez-Gamir et al., 2011; Ren et al., 2016; Wang et al., 2022; Adegoye et al., 2023). Additionally, we evaluated the genetic differences between E. humidus and common wheat (Triticum aestivum L.) using expressed sequence tag (EST) markers for wheat breeding. Barley EST markers were used to identify alien chromosomes in the wheat genetic background (Hagras et al., 2005). For wheat breeding programs, it is necessary to study wild species harboring environmental adaptations; therefore, we provided basic information on E. humidus observed in Korea when used as a useful genetic resource for improving wheat quality.
MATERIALS AND METHODS
Measurement of seed characteristics
To measure the seed length, width, height, and weight, 30 E. humidus seeds were randomly selected. Seed characteristics were measured using Vernier calipers. One hundred seeds were used to evaluate the germination ratio for one repetition, and the germination ratio was evaluated in triplicate. For statistical analysis, a t-test was performed to compare the differences in seed characteristics between E. humidus and common wheat (Triticum aestivum L. cv., Chinese Spring, CS).
Observation of mitotic chromosomes
Seeds of E. humidus were collected and germinated on wet filter paper in a Petri dish. After germination, the root tip was cut and placed in a fixation solution (3:1 ethanol:acetic acid, v/v) for at least five days at room temperature (25℃). Mitotic chromosomes were obtained from the meristem cells of the root tip using the squash method with 2% acetocarmine. The mitotic chromosomes of E. humidus were observed using a phase-contrast microscope equipped with a color camera (BX53F2 with DP-22, Olympus, Japan).
Observation of stomatal traits and leaf structure
Mature leaves of E. humidus and flag leaves of CS were selected to measure stomatal traits such as stomatal density (SD), guard cell length (GCL), and guard cell width (GCW) on the adaxial and abaxial sides. Stomata samples were obtained by placing nail polish on both sides of the leaf. The dried nail polish was removed from the surface of the leaves and placed on a slide to observe stomatal traits using a phase-contrast microscope with a color camera (BX53F2 with DP-22, Olympus, Japan). A t-test was performed for statistical analysis to compare the differences in stomatal traits between E. humidus and CS plants.
To investigate the leaf structure of E. humidus, leaf dissection was performed using a microtome (Minux S700 Rotary Microtome; RWD Life Science, China). Leaves of E. humidus were treated with a formalin-aceto-alcohol (FAA) fixation solution for at least one week. After fixation, the leaf samples were treated with 80% ethanol for 2 h, 90% ethanol for 2 h, and 99% ethanol for 2 h. Xylene was used to remove the ethanol, and the samples were placed in warm paraffin for 4 to 6 h for paraffinization. For slide preparation, the samples were dissected using a microtome, placed on slides in a floating bath, and dried on a slide warmer (XH-2016 Floating Bath & Slides Warmer, C&A Scientific, USA). The leaf structures of E. humidus and CS were observed using a phase-contrast microscope equipped with a color camera (BX53F2 with DP-22, Olympus, Japan).
PCR with EST markers
Barley and Leymus EST markers were used to identify polymorphisms in E. humidus and CS (Sato et al., 2003; Hagras et al., 2005; Bushman et al., 2008). The polymerase chain reaction (BIO-RAD T100™ Thermal Cycler, USA) was performed using the touchdown method. The conditions of touchdown method were modified with the method distributed by Hagras et al. 2005. The conditions were initial denaturation at 95℃ for 5 min, denaturation at 95℃ for 45 s, annealing at 65℃ to 60℃ (a decrease of 1℃ per one cycle) for 45 s, extension at 72℃ for 60 s in 5 sub-cycles, denaturation at 95℃ for 45 s, annealing at 60℃ for 45 s, extension at 72℃ for 60 s in 31 main sub-cycles, and then a final extension at 72℃ for 10 min. The PCR reaction mixture comprised 20 ng/μL of template DNA, 10 pmol/μL of a primer set, and 2X premix (Genet Bio, Korea) in a final volume of 25 µL. After the PCR, the amplicons were identified on a 1.5% agarose gel.
RESULT AND DISCUSSION
Seed characteristics of E. humidus
Seed size affects potential yield factors in wheat and barley, such as test weight and thousand grain weight (Spilde, 2013). The seeds of E. humidus were of the long-grain type, and the seed coat was dark brown (Fig. 1A). In addition, a purple color emerged from the seed coat of E. humidus after imbibition (Fig. 1B). E. humidus was smaller than that of common wheat (Triticum aestivum L. cv. Chinese Spring (CS); Table 1). The seed length of E. humidus (5.20 ± 0.25 mm) was approximately 85.67% of that of CS (6.07 ± 0.21 mm) and significantly different between them (p < 0.001). The seed width of E. humidus (1.38 ± 0.12 mm) was less than half that of CS (3.05 ± 0.15 mm) and was significantly different between them (p < 0.001). The seed height of E. humidus (0.90 ± 0.11 mm) was approximately 33.8% of that of CS (2.66 ± 0.14 mm) and significantly different between them (p < 0.001). Seed width and height differed significantly between E. humidus and the CS. For these reasons, the seed weight of E. humidus was approximately 12.8% of that of CS (p < 0.001). The seed size is an indicator of physiological quality and is related to germination, shoot length, radicle length, and shoot and root dry matter (Steiner et al., 2019). E. humidus initially showed the emergence of a coleoptile. Then the main root elongated like common wheat (Fig. 2A). The main roots of the germinated seeds were the rootlets (Figs. 2B and 2C). A comparison of seed germination ratios (Table 1) showed that E. humidus had a low germination ratio (6.0 ± 2.9%) compared to that of CS (85.7 ± 1.2%).
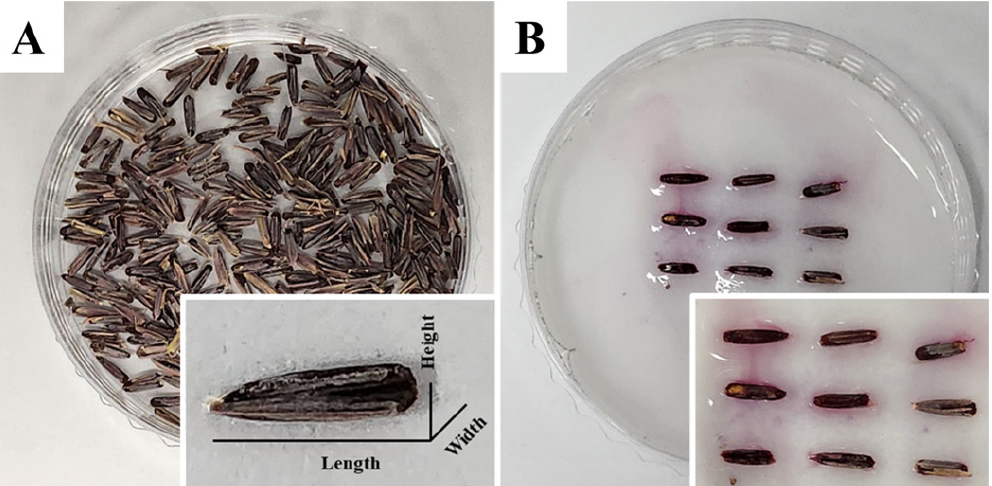
Table 1.
Evaluation of seed characteristics of Elymus humidus compared to common wheat (Triticum aestivum L. cv. Chinese Spring).
|
Seed characteristics Mean ± SD (n = 30) |
Length (mm) |
Width (mm) |
Height (mm) |
Weightz (g) |
Germinationy (%) |
| Elymus humidus | 5.20 ± 0.25*** | 1.38 ± 0.12*** | 0.90 ± 0.11*** | 0.11 ± 0.01*** | 6.0 ± 2.9*** |
| Chinese Spring | 6.07 ± 0.21 | 3.05 ± 0.15 | 2.66 ± 0.14 | 0.86 ± 0.01 | 85.7 ± 1.2 |

Leaf structure and stomatal traits of E. humidus compared to common wheat
We performed an anatomical dissection to investigate the leaf structure of E. humidus and compared it with that of the CS (Fig. 3). E. humidus showed a similar leaf structure to that of common wheat, i.e., the xylem was observed in the middle of the vascular bundle in the main vein, and there were small vascular bundles on both sides of the main vein (Fig. 3). On average, the ratio of the main vascular bundle diameter to the main vein thickness of E. humidus was lower than that of common wheat (p < 0.05). In contrast, the ratio of the minor vascular bundle diameter to the main vein thickness of E. humidus was not significantly different from that of CS (Table 2). The xylem dimensions are important for leaf water transport (Scoffoni et al., 2017). In the main vascular bundle, E. humidus showed a smaller xylem than CS (p < 0.001) and no difference in phloem size compared to common wheat (Table 2). In addition, E. humidus showed a thinner epidermis (p < 0.001), and the sclerenchymatous bundle sheath (p < 0.01) was significantly thicker in the CS.

Table 2.
Evaluation of stem characteristics of Elymus humidus compared to common wheat (Chinese Spring) by microdissection with a microtome.
| Variety |
Main vascular bundle diameter/ Main vein thickness |
Minor vascular bundle diameter/ Main vein thickness | Main vascular bundle (μm) |
Epidermis thickness (μm) |
Sclerenchymatous bundle sheath (μm) | |
| Xylem | Phloem | |||||
| Elymus humidus | 3.26 ± 0.30* | 2.76 ± 0.33 | 21.61 ± 3.19*** | 7.37 ± 1.53 | 21.76 ± 2.82*** | 24.09 ± 4.61** |
| Chinese Spring | 4.26 ± 0.19 | 2.66 ± 0.27 | 28.88 ± 2.01 | 8.40 ± 1.69 | 28.42 ± 2.44 | 30.55 ± 3.77 |
Stomata play an important role in photosynthesis and are correlated with the chlorophyll content index, which is correlated with yield potential (Jones, 1998; Rahim et al., 2021). In addition, stomatal morphology varies according to different plant types and is related to water efficiency and gas exchange (Fryns-Claessens & Van Cotthem, 1973; Nunes et al., 2020). According to Fryns-Claessens & Van Cotthem (1973) and Nunes et al. (2020), the stomata on the adaxial and abaxial sides of E. humidus showed a graminoid type similar to that of common wheat; however, the stomatal arrangement of E. humidus was different from that of common wheat (Fig. 4). Hence, stomatal traits, such as length, width, density, and distance between stomata, were evaluated (Table 3). Size of stomata is related to gas exchange, i.e., as carbon dioxide increases in wheat, the size of stomata is small and their density is high (Jordan et al., 2020). On the adaxial side, the stomatal traits of E. humidus were significantly different from those of common species. The length, width, and density of E. humidus stomata were approximately 93.6%, 84.1%, and 80.2%, respectively, of those of common wheat. In contrast, the distance between the stomata of E. humidus was approximately 1.53-times greater than that of common wheat. The abaxial side of E. humidus exhibited a tendency similar to that of the adaxial side. The length, width, and density of E. humidus stomata were approximately 82.7%, 72.7%, and 67.8% of those of common wheat, respectively. The distance between the stomata of E. humidus is approximately 1.78-fold greater than that of common wheat. To improve wheat quality, it is necessary to evaluate stomatal traits and capacity for gas exchange in future cross lines of common wheat and E. humidus.

Table 3.
Evaluation of stomatal traits of Elymus humidus compared to common wheat (Chinese Spring).
| Stomatal traits | Adaxial side (Mean ± SD, µm) | |||
| Density | Length | Width | Distance | |
| Elymus humidus | 49.33 ± 3.21*** | 46.37 ± 2.63*** | 23.59 ± 1.83*** | 140.43 ± 16.95*** |
| Chinese Spring | 61.50 ± 7.58 | 49.56 ± 3.21 | 28.04 ± 3.41 | 91.84 ± 14.39 |
| Stomatal traits | Abaxial side (Mean ± SD, µm) | |||
| Density | Length | Width | Distance | |
| Elymus humidus | 29.00 ± 2.00*** | 46.63 ± 2.78*** | 20.02 ± 1.48*** | 249.08 ± 38.51*** |
| Chinese Spring | 42.80 ± 3.85 | 56.40 ± 4.26 | 27.54 ± 1.82 | 140.17 ± 22.83 |
Identification of chromosomes and polymorphism of E. humidus versus common wheat
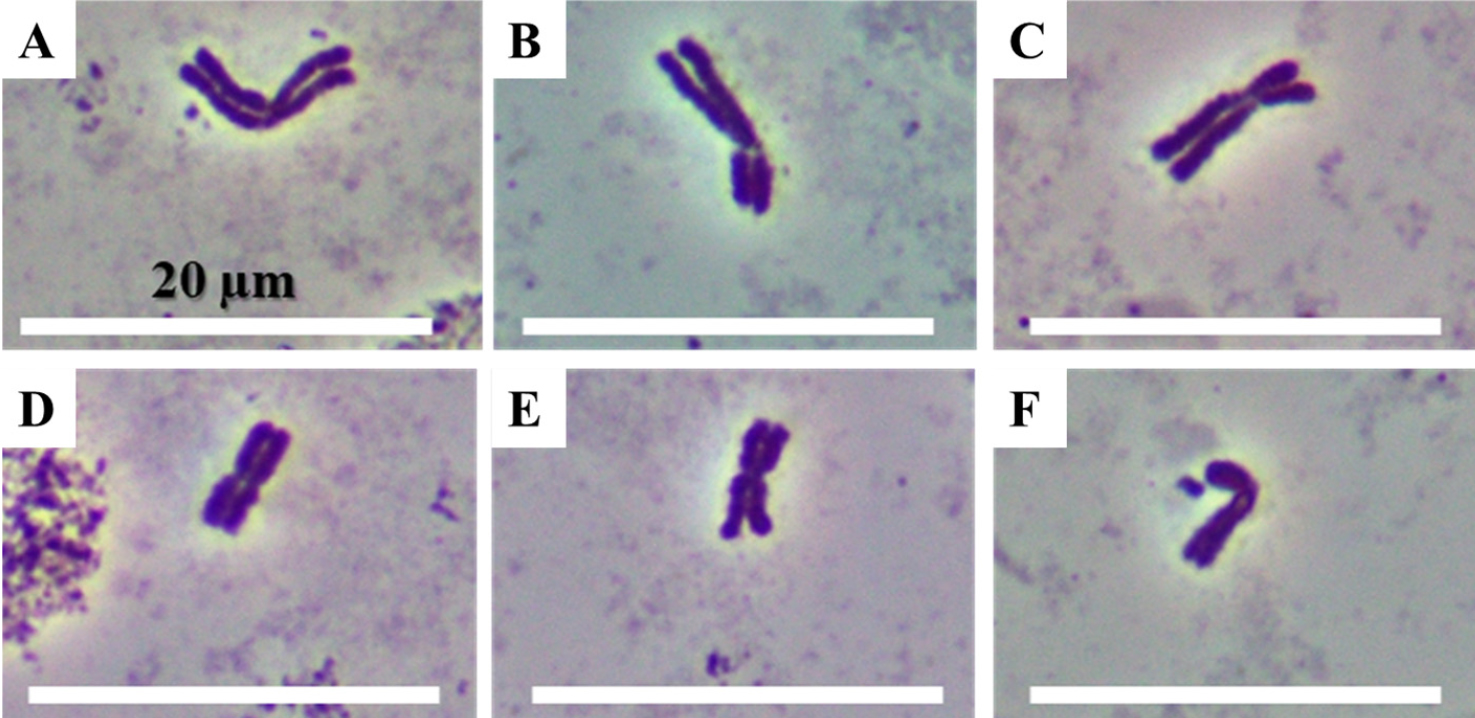
E. humidus (2n = 6x) in Japan has 42 chromosomes, and the number of chromosomes was the same as that of common wheat (Muramatsu et al., 1993). We observed chromosomes of E. humidus founded in Korea (Fig. 5). Chromosome karyotype is one of methods for identification of chromosome e.g. relative chromosome length, ratio of short arm of long arm, and with micro-satellite or not (Gill et al., 1991). E. humidus showed long and short chromosome relatively (Fig. 6). In the chromosomes with relative long length, the ratio of short arm to long arm was approximately 1 to 1 or 1 to 2 (Fig. 6A-C). In the chromosomes with relative short length, the ratio of short arm to long arm was approximately 1 to 1 and 1 to 1.5 (Fig. 6D-F). Also, some chromosomes with relative short length were with micro-satellite (Fig. 6F).
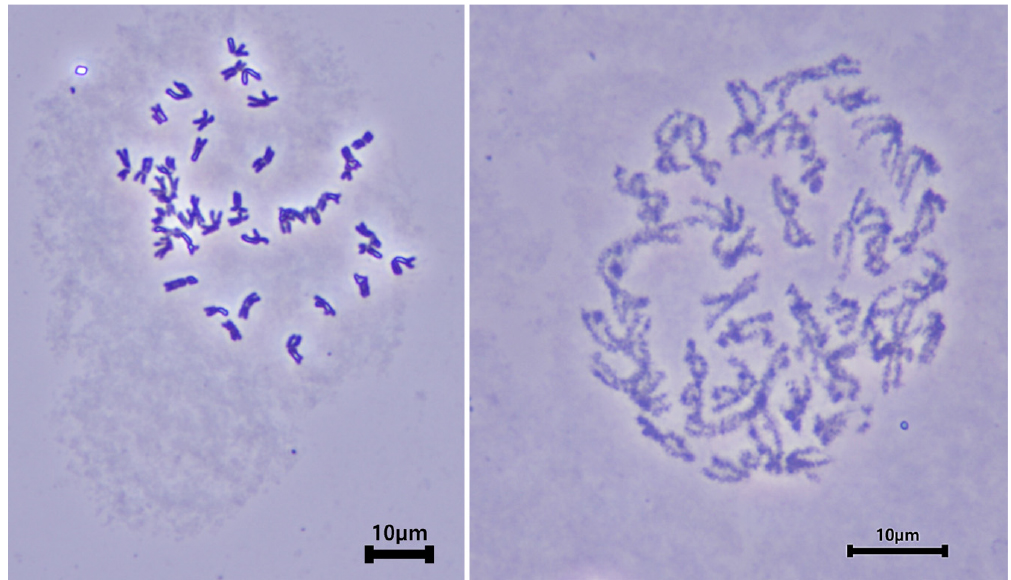
DNA markers are used to identify target genes or alien chromosomes in the genetic background of wheat (Hagras et al., 2005; Du et al., 2022). Hagras et al. (2005) evaluated Triticeae species, including wild species, using barley EST markers, and Bushman et al. (2008) developed Leymus EST. We used 236 EST markers (124 barley and 112 Leymus EST markers) to evaluate genetic diversity (Fig. 7). Among the 236 markers, there were 67 markers (14 from barley and 53 from Leymus) for monomorphisms between E. humidus and CS, 12 markers (2 from barley and 10 from Leymus) for CS only, and 58 markers (51 from barley and 7 from Leymus) for E. humidus only. Finally, 38 EST markers (16 from barley and 22 from Leymus) with polymorphisms in E. humidus and CS were identified. No other 61 markers (41 from barley and 20 from Leymus) were used to identify E. humidus or CS. These markers are expected to be useful in wheat breeding programs using E. humidus as a genetic resource.

In conclusion, the increasing influence of climate change has a negative effect on crop growth (Shew et al., 2020). In this situation, it is necessary to consider the accessibility of wild species with strong environmental adaptability and their agronomic value when using wild plants. Hence, in this study, we investigated the biological characteristics of E. humidus because it shows tolerance to excess moisture (Sasanuma et al., 2002). E. humidus had small seeds compared to common wheat and a low germination ratio; it had small xylems and epidermal thicknesses and a leaf structure similar to that of common wheat. Unfortunately, we investigated only the leaf structure in this study because collecting root samples in the field is difficult and E. humidus, a wild species, showed a low germination ratio. Nevertheless, observation of the root structure is necessary to verify the correlation between root structure and excess moisture tolerance. E. humidus had 42 chromosomes and a structure similar to that of common wheat. For interspecific crossing in the wheat genetic background, studying chromosomes in more detail is necessary. However, barley EST markers allowed the detection of polymorphisms between E. humidus and common wheat.
