

서 론
재료 및 방법
시험재료 재배 및 환경
종실의 일반성분, 총 식이섬유, sucrose 분석
총 폴리페놀과 플라보노이드 함량
화분 활력 및 주사전자현미경 검정
통계분석
결과 및 고찰
온도구배온실 온도
상승된 온도에 따른 팥 생육 및 수량 변화
팥의 일반성분 비교
팥의 에탄올 추출물의 항산화성분의 함량
생육시기에 따른 고온 스트레스시 화분 형태와 활력 비교
적 요
서 론
팥(Vigna angularis L.)은 일년생 콩과 식물로 한국을 비롯한 온대 동북 아시아 지역에서 주로 재배되어 왔다(Rho et al., 2003; Song et al., 2016). 2020년 한국의 팥 재배면적은 4,931 ha로 5,809 ton이 생산되었으며 생산물은 주로 제과 제빵용 앙금으로 사용되며 두과 작물에서 콩 다음으로 재배면적이 많은 작물이다(MAFRARK, 2021).
팥은 콩에 비하여 수량은 낮지만 토양 및 기후적응성이 좋아 작부체계에 유용하게 이용될 수 있다(Song et al., 2016). 개화시기가 가뭄 스트레스에 취약한 것으로 알려져 기후변화에 따른 수량의 변동성이 심한 작물이다(Han et al., 2021). 팥의 전생육기 생육 적온은 주야간 평균 20°C으로 알려져 있으며, 개화기 적온은 24°C이고 16°C 이하로 내려가면 화아 분화에 영향을 미쳐 화아 생성에 불량한 환경을 조성하여 협수가 줄어든다(RDA, 2018).
최근 10년(2011 - 2021년) 지구 지표 평균 온도는 과거 10년(1850 - 1900년)보다 약 0.99°C 상승하였으며 배출된 CO2양도 관측 이래 최고 수준이고, 100년간 우리나라 평균 기온이 약 1.8°C 상승 하였으며 이는 세계 평균 보다 높은 수치로 극한 기온 및 이상 기후의 발생 빈도를 높인다는 보고가 있다(IPCC, 2022). 농업은 기후 환경에 매우 민감한 산업으로 그 중 온도는 작물의 생리 및 생육 반응과 수량 및 품질에 영향을 미쳐 기후변화에 의한 온도의 상승은 전세계 식량안보와 연관 되어있고 작물의 성장, 대사 작용, 수량을 제한하는 주요 요인이다(Bita & Gerats, 2013; Hasanuzzaman et al., 2013; Wheeler & Von Braun, 2013). 여러 작물에서 고온 스트레스에 대한 연구가 수행되었다. 콩(Glycine max L.)은 고온 조건에서 협수와 종실 무게가 변해 품종에 따라 수량이 증가하거나 감소했으며(Lee et al., 2018), 35°C이상의 고온에서는 수량의 27%가 감소했다(Gibson & Mullen, 1996; Puteh et al., 2013). 또한 고온은 콩의 영양생장기의 생장에 긍정적인 효과를 주었지만 개화기부터 수량과 밀접한 영향을 받아 착협기에서 종실비대기까지 환경에 지배적인 영향을 준다고 보고되었다(Kantolic et al., 2013). 옥수수(Zea mays L.)는 온도가 상승함에 따라 생육발달이 빨라져 각 생육시기의 기간이 짧아지며 고온 스트레스 하에서 꽃가루 생존력이 감소해 수량이 감소하였고(Lizaso et al., 2018), 밀 (Triticum aestivum L.)의 경우 CO2 농도가 대조구 대비 100 ppm으로 증가한 조건에서도 고온에 의한 수량감소는 10 - 12%로 피해가 회복되지 못했다(Cai et al., 2016). 팥 종자에는 천연 항산화성분이 많이 포함되어 있다. 이에 대해 항산화 성분 추출 및 특성 분석법에 관해 연구가 발표되었고(Randhir et al., 2004), 품종별로 이화학적 특성 및 항산화 활성이 다르다고 알려져 있다(Woo et al., 2010; Sung et al., 2020). 연차별 기상상황과 과습 토양에 따른 노지 재배환경 조건 변화로 팥 품종별 sucrose, protein, 폴리페놀(polyphenol)과 플라보노이드(flavonoid)함량 등이 유의한 차이를 보이는데(Chun et al., 2017; Oh et al., 2022), 빈번한 고온 영향이 팥의 항산화 성분의 변화에 어떤 영향을 미치는지는 자세히 보고된 바가 없다. 빈번한 기후변화에 대한 수확량 변동 예측 모형 개발, 환경 제어 챔버에서 고온 스트레스 연구가 다양한 작물에 대한 고온 피해 연구가 콩, 배추, 벼 등 다양한 작물 연구에서 이뤄지는 것에 비해(Lee et al., 2015; Lee et al., 2018; Oh et al., 2018; Poter & Semenov, 2005) 팥에 대한 환경 스트레스 연구는 토양수분 스트레스(Chun et al., 2017)와 연차별 노지재배시 강수량 변화에 대한 팥의 수량 및 항산화성분 변화(Oh et al., 2022)외에 고온 영향 연구는 보고되지 않아 고온 환경에서 팥의 생육 및 생리적 변화에 대한 연구가 매우 필요하다.
본 연구에서는 지구기후변화에 대비하여 생육시기별 고온 및 습도 영향이 팥의 수량에 영향을 미치는지를 살펴보고자 하였다. 팥의 고온 피해 연구를 수행하기 위해 환경 조건이 노지와 유사하나 고온 조성이 가능한 반폐쇄 형태의 온도구배온실을 고안하여 국립식량과학원 남부작물부 내에 설치하여 연구하였다. 따라서 노지와 유사한 환경의 단계별 온도 및 습도 변화 환경에 따라 현재 팥 재배 품종 중 도복이 강하고 내재해성과 앙금가공적성이 우수하고 기계수확이 가능하여 현재 가장 많이 재배되고 있는 품종으로 ‘아라리’(Song et al., 2016)를 사용하여 생육시기별 적온 데이터를 축적하고 온도 및 습도 변화가 화분 임성과 수량, 종실일반성분 및 항산화성분에 미치는 영향을 분석하여 팥의 고온 재배시 기초자료를 제공하고자 수행하였다.
재료 및 방법
시험재료 재배 및 환경
본 연구는 2021과 2022년 밀양시 내이동에 위치한 국립식량과학원 남부작물부 실험 포장(N35° 29′ 39.1″ E 128° 44′ 24.5″)내 노지, 비가림온실, 온도구배온실에서 수행하였다. 비가림온실은 가로 8 m × 세로 30 m × 높이 6 m의 크기로 측면 비닐은 강우시를 제외하면 올라가 있는 앞뒤의 비닐이 설치되지 않은 노지와 기온이 비슷한 온실이다. 온도구배온실은 가로 2.4 m × 세로 30 m × 높이 2.5 m로 한면은 뚫려있고 한면은 폐쇄된 형태로 배기팬이 온실 내부 공기를 한 방향으로 배기하여 점진적인 온도구배가 형성되도록 제작하였다. 온도구배온실 내의 시험구는 일정 너비로 4등분(T1 < T2 < T3 < T4)하여 구분하였다(Fig. 1). 온도구배온실 폐쇄 면에 배기팬을 지름 50 cm 2개, 25 cm 4개를 설치하여 내부 공기 흐름 및 온도 구배를 조절하였다(Fig. 1). 온도구배온실 각 처리구 정중앙에 온/습도 센서를 지표 40 cm 높이로 설치하였으며 온도를 1분단위로 측정해 배기팬의 가동여부 및 풍력을 조절하여 온도구배를 형성하였다. 구배 설정 및 환경 데이터 수집을 위하여 비가림하우스와 노지 지표 1.5 m 높이에 온도구배 온실과 동일한 온/습도 센서(J.EZ-020, Jnp system Inc., Korea)를 설치하여 10분 간격으로 수집하여 엑셀을 활용하여 분석하였다. 외부와 T4 처리의 온도 차이가 8°C일 시 50 cm 팬 두 개가, T1과 T4 처리의 온도 차이가 5°C 이상일 때 상부의 25 cm 팬 두 개가, T1과 T4 처리의 온도 차이가 3°C 이상일 때 하부의 25 cm 팬 두 개가 80% 풍력으로 가동되도록 설정하여 온도구배온실 내부온도가 외부 대비 5°C 이상 상승하지 않고 내부의 온도 구배가 외기 대비 4 - 5°C을 유지하도록 하였다(Fig. 1). 습도는 10분 단위로 측정해 온도구배온실 내 습도가 일정하게 유지 될 수 있도록 추가 관수 하였다. 야간은 약 4.2% 차이가 났으며 주간에는 최대 9.9% 차이를 나타났다(Fig. 2). 시험에 사용한 팥은 아라리(ARR)로 1주 2본 와그너 포트(1/2000) 직파하였으며 토양은 N-P2O5-K2O를 사용 10 a당 4.2-5.2-5.8 kg로 시비하여 표준재배법에 준하였다(RDA, 2018). 전생육기 실험군은 비가림온실에서 재배하였고 각 생육시기별 실험군은 비가림온실에서 재배하고 있다가 포트 이동하여 온도구배온실에서 고온 처리를 하였고 시기가 지나면 비가림온실로 옮겼다. 관수는 토양 수분이 20 - 30%를 유지하도록 수행하였다. 생육 조사는 경장, 경태, 주경절수, 분지수, 협수, 백립중, 수량을 조사하였다(RDA, 2018).
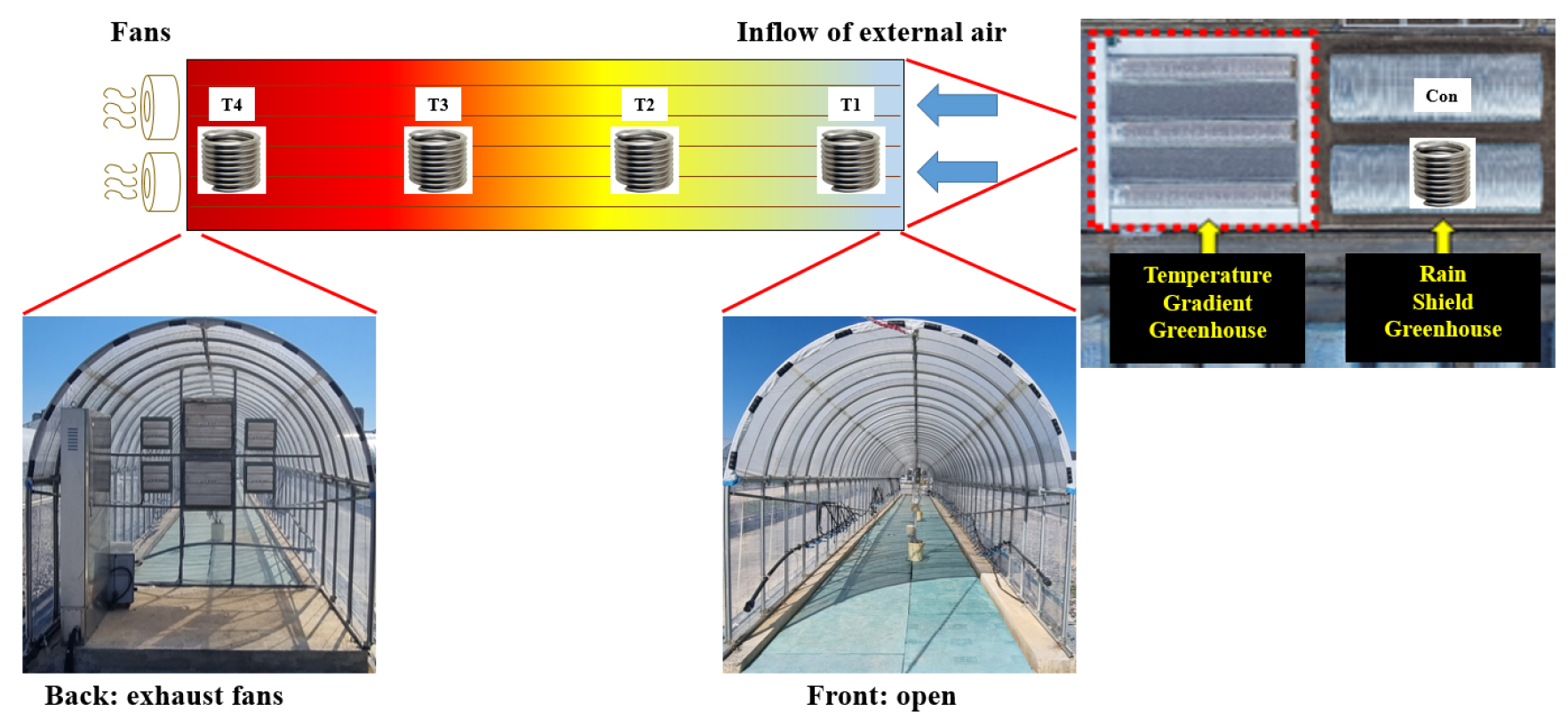
종실의 일반성분, 총 식이섬유, sucrose 분석
팥 종실의 일반성분 중 수분과 회분은 상압가열건조법, 단백질은 Micro-kjeldahl법, 총 식이섬유는 효소 중량법을 이용하여 분석하였다. 팥의 sucrose는 시료에 50% ethanol 50 ml를 넣어 추출한 분석용액을 reciprocal shaker로 200 rpm에서 15분 섞어준 후 80°C 항온수조에서 25분간 초음파 처리 후 0.2 μm syringe filter로 여과 한 후 HPLC (SI-2, SHISEIDO, Japan)을 사용하여 분석하였다. 분석을 위한 칼럼은 UK-Amino 250 x 3 mm, 유속은 0.4 ml/min하여 검출은 RI detector를 이용하였다.
총 폴리페놀과 플라보노이드 함량
시료의 항산화 성분 분석 하기 위하여 시료를 분쇄한 후 80% 에탄올로 추출 후 SHIMADZU UV-spectrophotometer UV-1900 (Shimadzu, Japan)을 이용하여 폴리페놀은 Singleton & Rossi (1965) 방법, 플라보노이드는 Zhishen et al. (1999) 방법을 따라 흡광도법으로 분석하였다.
화분 활력 및 주사전자현미경 검정
전생육기의 개화한 팥의 식물체 당 3개 이상의 화기를 모아서 화분 활력 및 형태를 측정하였다. 실험군 개체별 화분을 슬라이드 글라스에 분리하여 alexander 염색용액(Peterson et al., 2010)으로 10분 간 염색한 뒤 실체현미경(Leica M125, Germany)으로 화분 및 화분 활력을 조사하였다. 주사전자현미경(Scanning electron microscopy)을 이용하여 시료를 촬영 하기 위해서 화분을 건조시키고 알루미늄 stub에 절연테이프를 붙여서 화분을 고정시킨다. 화분 표면 코팅은 sputter coater (Hitachi, Japan)을 이용하여 60초 12 mA로 백금 10 mÅ 두께로 하였다. SEM (Hitachi, Japan)을 이용하여 화분 표면 이미지를 얻었다.
통계분석
조사된 데이터는 SAS 9.4 (SAS Institute, Cary, NC, USA)을 사용하여 ANOVA (P<0.05)분석을 통하여 유의성 검정을 하였다.
결과 및 고찰
온도구배온실 온도
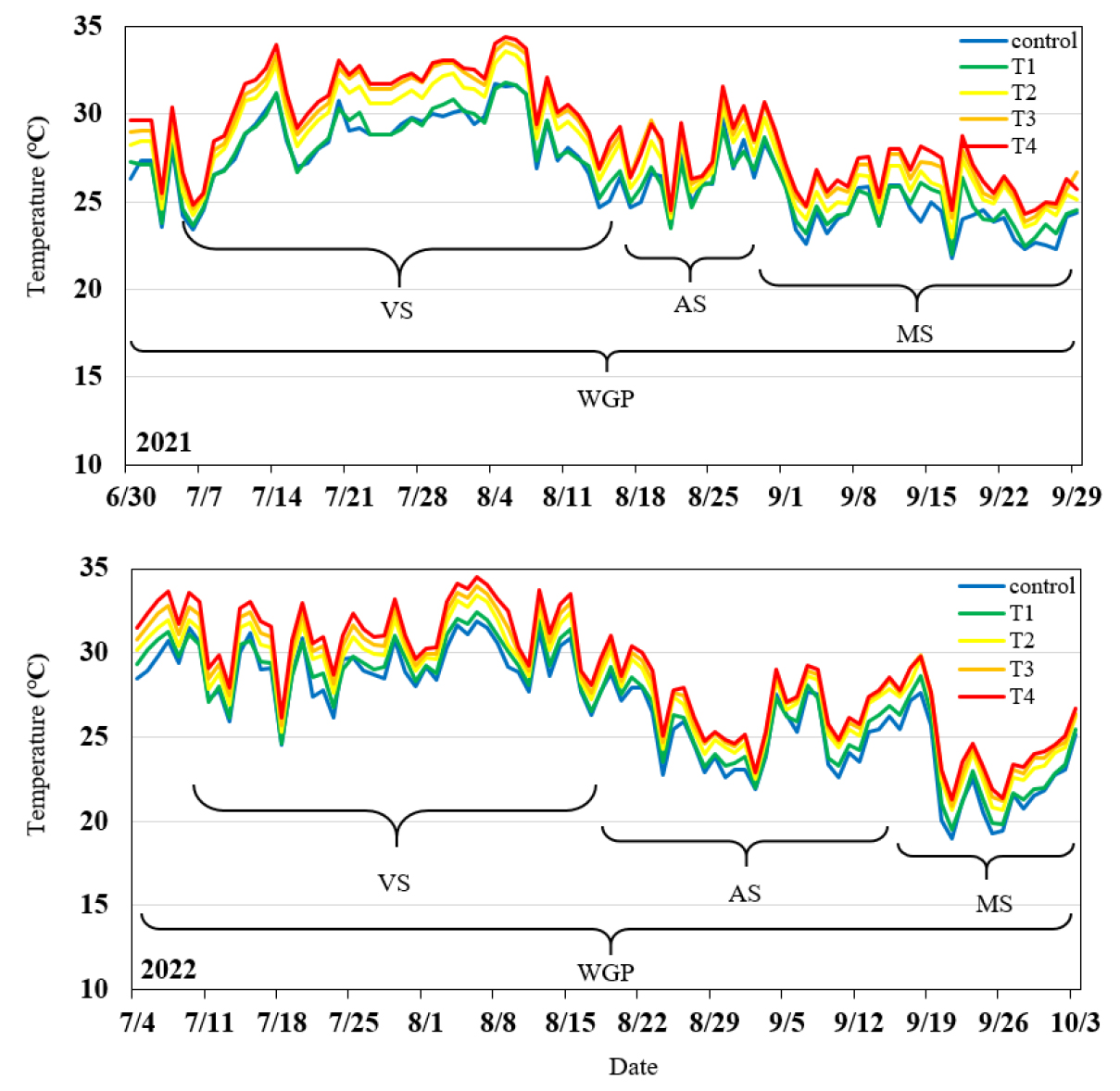
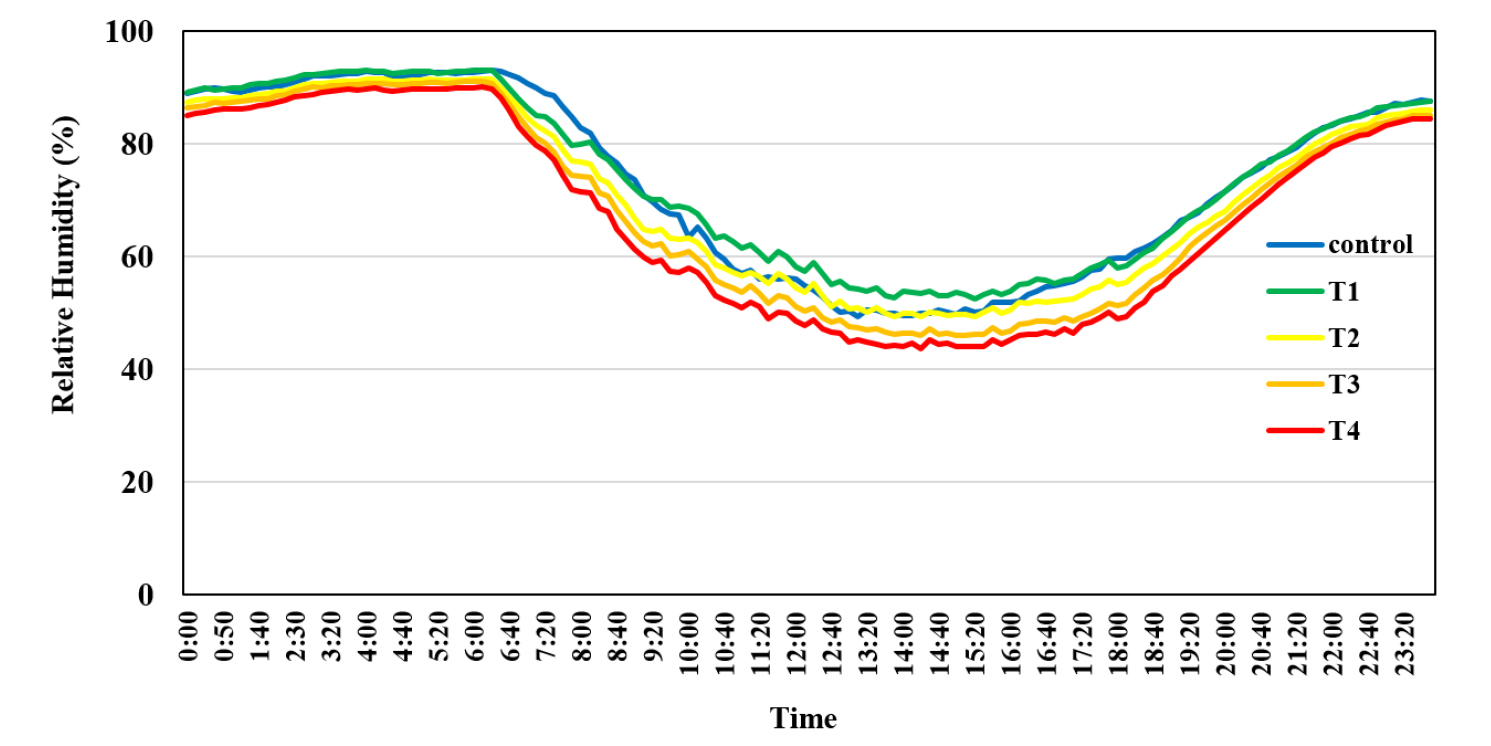
온도구배온실은 내부를 균등하게 4구역으로 나누어 T1 - T4으로 나누어 일평균 변화를 확인하였다(Figs. 1 and 2). 습도는 시간대 별 평균 변화가 주간에는 고온의 영향으로 내부 온도가 상승할수록 환풍기 가속 영향이 있어 최저 43.7 (T4) - 50.7 (Control)%였고 야간에는 85.1 (T4) - 93.5 (Control)%였다(Fig. 3). 옥수수의 노지 온실 고온 스트레스 실험 시 유지하였던 환경 변화 온도는 최저 19°C, 최고 44.5°C였고, 습도는 30 - 80%였으며 주간은 습도가 40%까지 떨어졌다(Lizaso et al., 2018). 이 실험 환경보다 밀양의 온도구배온실에서 습도 변화가 구획별로 일정하게 유지되면서 습도 및 고온 스트레스 영향으로 팥이 재배 되었다. 팥은 유묘기, 신장기, 개화기, 협신장기 및 등숙기 등 생육단계를 5개로 나눌 수 있다(RDA, 2018). 세분된 단계는 실험상 그 구별이 어려워 유묘기와 신장기를 영양생장기로 합치고, 개화기, 협신장기와 등숙기를 등숙기로 합쳐 세개의 생육 단계와 전생육기로 나눠서 실험하였다. 영양생장기에 화아 형성 시기까지 포함하였고 개화기는 화아가 열리는 시기부터 협이 맺혀 화아가 떨어지는 시기로 한정하였다.
생육시기별은 2021년 전생육기(6월 30일 - 9월29일; 92일), 2022년 전생육기(7월 4일 - 10월 3일; 92일)이고 2021년 영양생장기(7월 7일 - 8월 17일; 42일), 2022년 영양생장기(7월 11일 - 8월 18일; 39일), 2021년 개화기(8월 18일 - 8월 30일; 13일), 2022년 개화기(8월 19일 - 9월 14일; 27일), 2021년 등숙기(8월 31일 - 9월 29일; 30일), 2022년 등숙기(9월 15일 - 10월 3일; 19일)이었다. 개화시기는 연차별 차이는 있었지만 온도 상승과 차이를 보이지 않아 같은 시기에 포트 이동을 하였다. T1와 T4의 온도 차이는 3.5 – 4.0°C로 온도 구배 형성을 확인하였으며, 팥 생육 기간 중 고온에 의한 피해 연구 환경 조건이 조성되었음을 확인하였다. 온도 구배 지속시간은 일사가 유지되는 7시부터 끝나는 19시까지 약 12시간이었다. 2021와 2022년 최저, 평균, 최고 기온 연차 간 차이를 보면 전생육기에서는 0 - 1°C 정도 차이가 있었고, 영양생장기에서는 0.4 - 3.7°C로 T2처리에서 최고 온도일 때 2021년이 약 3.7°C 높았다. 2022년 개화기는 최저 온도가 2021년 보다 약 2.0 - 2.4°C 낮아져 협의 생성이 늦어져 13일에서 27일로 14일 더 개화기가 지속되었다. 등숙기는 0 - 3.9°C 정도 차이를 보이고 T2처리에서 최고 온도 일 때 2022년이 3.9°C 높았다. 하지만 2021년보다 2022년 등숙기의 최저온도가 1.6 - 1.9°C 낮고 최고온도는 1.7 - 3.9°C로 더 높아 온도 편차는 2022년이 높았다(Table 1).
Table 1.
Comparison of temperature low, average, and high temperatures (°C) of whole growth period (WGP), vegetative stage (VS), anthesis stage (AS), and mature stage (MS) in 2021 and 2022.
| Treatmentz | Air temperature (°C) | ||||||||||||
| WGP | VS | AS | MS | ||||||||||
| 2021 | 2022 | Dy | 2021 | 2022 | D | 2021 | 2022 | D | 2021 | 2022 | D | ||
| Con | L | 21.3 | 21.7 | -0.4 | 22.8 | 23.4 | -0.6 | 22.5 | 20.4 | 2.1 | 19.4 | 17.7 | 1.7 |
| A | 26.6 | 26.6 | 0.0 | 28.6 | 27.6 | 1.0 | 26.7 | 25.1 | 1.6 | 24.1 | 24.0 | 0.1 | |
| H | 33.3 | 34.3 | -1.0 | 35.5 | 34.4 | 1.1 | 32.6 | 31.9 | 0.7 | 30.9 | 33.9 | -3.0 | |
| T1 | L | 22.2 | 22.1 | 0.1 | 23.7 | 23.7 | 0.0 | 22.8 | 20.8 | 2.0 | 20.0 | 18.1 | 1.9 |
| A | 26.9 | 27.0 | -0.1 | 28.7 | 28.0 | 0.7 | 26.6 | 25.5 | 1.1 | 24.5 | 24.5 | 0.0 | |
| H | 33.9 | 33.6 | 0.3 | 36.4 | 33.8 | 2.6 | 32.4 | 31.4 | 1.0 | 31.3 | 33.0 | -1.7 | |
| T2 | L | 23.0 | 22.6 | 0.4 | 24.6 | 24.2 | 0.4 | 23.5 | 21.1 | 2.4 | 20.6 | 18.8 | 1.8 |
| A | 28.1 | 27.9 | 0.2 | 30.2 | 28.8 | 1.4 | 27.6 | 26.3 | 1.3 | 25.5 | 25.7 | -0.2 | |
| H | 35.9 | 35.3 | 0.6 | 38.7 | 35.0 | 3.7 | 34.3 | 33.6 | 0.7 | 31.3 | 35.2 | -3.9 | |
| T3 | L | 23.1 | 22.8 | 0.3 | 24.9 | 24.3 | 0.6 | 23.6 | 21.4 | 2.2 | 20.7 | 19.1 | 1.6 |
| A | 28.7 | 28.4 | 0.3 | 30.8 | 29.3 | 1.5 | 28.3 | 26.7 | 1.6 | 26.1 | 26.1 | 0.0 | |
| H | 35.9 | 35.9 | 0.0 | 38.7 | 36.0 | 2.7 | 34.3 | 33.9 | 0.4 | 32.9 | 35.3 | -2.4 | |
| T4 | L | 23.1 | 22.9 | 0.2 | 24.7 | 25.2 | -0.5 | 23.6 | 21.4 | 2.2 | 20.7 | 19.0 | 1.7 |
| A | 29.0 | 28.8 | 0.2 | 31.2 | 30.5 | 0.7 | 28.5 | 27.0 | 1.5 | 26.4 | 26.4 | 0.0 | |
| H | 37.6 | 36.9 | 0.7 | 40.3 | 37.9 | 2.4 | 35.8 | 34.6 | 1.2 | 34.7 | 36.4 | -1.7 | |
상승된 온도에 따른 팥 생육 및 수량 변화
온도 처리에 따른 생육 특성 조사한 결과는 Table 2와 같다. 온도가 상승하더라도 ‘아라리’의 경장, 경태, 마디수, 분지수는 대조구와 고온처리구에서 유의미하게 변하지 않았다. 2021년 경장은 전생육기 T2 (최저온도 23.0°C; 평균온도 28.1°C; 최고온도 35.9°C)에서 가장 크고 55.1 ± 6.4 cm로 가장 크고, 영양생장기 T3 (최저온도 24.9°C; 평균온도 30.2°C; 최고온도 38.7°C)에서 36.4 ± 3.0 cm로 가장 낮은 값을 나타냈으며 영양생장기와 개화기 외 다른 생육시기에서는 T4에서 가장 낮은 값을 나타냈다. 2022년은 경장은 등숙기 T2 (최저온도 18.8°C; 평균온도 25.7°C; 최고온도 35.2°C)에서 가장 높은 값인 58.8 ± 5.6 cm, T1 (최저온도 18.1°C; 평균온도 24.5°C; 최고온도 33.0°C)에서 가장 작은 값인 47.2 ± 3.5 cm였다. 연차별 ‘아라리’ 생육 특성은 최저 최고값의 범위가 다르고 연간 온도에 변화에 따라 다르고 차이가 뚜렷하게 보이지는 않았다. 하지만 2021년 각 생육시기별 온도가 높아질수록 경태와 경장은 낮은 값을 유의미있게 보여줬지만 2022년 분지수에서는 각 처리별 온도별 차이가 크지 않은 것으로 나타났다. 2021년 경장은 전생육기를 제외한 T4 처리에서 낮은 값은 보였다. 반면 2022년 경장은 전년 대비 대조구보다 증가하고 고온일수록 유의미하게 증가하였다. 수분 스트레스를 받은 팥의 다양한 품종에서 경장, 경태, 마디수, 분지수가 낮아지는 것으로 보아 그에 유사한 스트레스를 받은 것으로 유추할 수 있었다 (Chun et al., 2017).
Table 2.
Growth components of “Arari” grown to high temperatures in 2021 and 2022.
| Year | Treatmentz | Stem length (cm) | Stem diameter (mm) | No. of nodes | No. of branches | |
| 2021 | Con | 38.7±4.7ghiy | 7.1±1.0cde | 13.7±1.8bcdef | 2.0±0.8bcd | |
| WGP | T1 | 51.9±6.8ab | 7.4±0.7cd | 15.0±1.6abc | 2.8±0.9abc | |
| T2 | 55.1±6.4a | 7.2±0.4cde | 14.5±1.5abcd | 2.6±0.5abc | ||
| T3 | 51.7±6.6ab | 6.6±0.6cdef | 15.3±0.8a | 2.9±1.0ab | ||
| T4 | 48.2±7.7bc | 6.2±0.9ef | 14.3±0.8abcd | 2.7±1.2abc | ||
| VS | T1 | 45.2±7.7cdef | 8.8±2.2a | 14.0±1.6abcde | 3.5±1.4a | |
| T2 | 37.2±6.3hi | 6.8±2.0cdef | 13.2±0.9def | 2.0±0.9bcd | ||
| T3 | 36.4±3.0i | 7.6±1.4abc | 13.5±1.8cdef | 2.2±1.1bcd | ||
| T4 | 39.2±3.5ghi | 8.4±1.3ab | 13.7±1.3bcdef | 2.2±0.6bcd | ||
| AS | T1 | 47.5±2.9bcd | 6.2±0.5ef | 11.5±1.3g | 1.8±0.6cd | |
| T2 | 43.7±3.2cdefg | 6.2±0.5ef | 12.5±1.7fg | 2.3±0.8bcd | ||
| T3 | 42.8±3.5defg | 6.2±0.8ef | 12.4±1.4fg | 1.5±0.5d | ||
| T4 | 44.6±2.7cdef | 6.4±0.6def | 13.8±1.4bcdef | 2.8±0.9abc | ||
| MS | T1 | 42.1±2.9efgh | 6.7±0.7cdef | 14.9±1.2abc | 2.4±1.1bcd | |
| T2 | 46.2±4.4cde | 6.3±0.6def | 15.2±0.4ab | 1.8±1.1cd | ||
| T3 | 46.7±4.3cde | 7.1±0.9cde | 15.4±1.8a | 2.3±1.3bcd | ||
| T4 | 40.7±5.0fghi | 5.7±1.1f | 12.8±2.1efg | 2.3±1.1bcd | ||
| 2022 | Con | 54.2±4.6abcd | 7.1±0.4abc | 15.9±0.9ab | 3.6±1.4ab | |
| WGP | T1 | 53.4±5.0bcde | 7.4±0.8a | 15.1±0.7abc | 3.7±1.1ab | |
| T2 | 57.4±2.5ab | 6.6±0.6bcd | 15.5±0.5abc | 3.0±0.8b | ||
| T3 | 57.4±4.4ab | 6.8±0.6abc | 15.9±1.1ab | 4.0±0.5ab | ||
| T4 | 57.2±4.7ab | 6.6±0.7bcd | 15.6±1.3abc | 4.4±1.0a | ||
| VS | T1 | 52.1±6.7cde | 7.1±1.2abc | 14.6±1.2c | 4.0±1.6ab | |
| T2 | 56.1±3.5ab | 6.4±0.5cd | 15.0±0.8bc | 4.0±1.1ab | ||
| T3 | 55.3±3.1ab | 6.4±0.7cd | 15.2±1.3abc | 3.8±0.8ab | ||
| T4 | 57.2±3.5ab | 6.0±0.9d | 15.4±1.1abc | 3.8±1.5ab | ||
| AS | T1 | 50.1±4.2def | 7.1±0.4abc | 15.6±0.5abc | 3.6±1.5ab | |
| T2 | 53.6±4.8bcde | 7.1±0.6abc | 15.9±1.3ab | 3.3±1.1ab | ||
| T3 | 52.9±4.2bcde | 6.9±0.7abc | 15.8±0.9ab | 3.3±0.9ab | ||
| T4 | 49.4±4.2ef | 7.1±0.9abc | 15.2±0.8abc | 3.8±1.0ab | ||
| MS | T1 | 47.2±3.5f | 7.3±0.8ab | 16.2±1.1a | 4.0±1.3ab | |
| T2 | 58.8±5.6a | 7.0±0.5abc | 15.9±1.0ab | 3.3±0.5ab | ||
| T3 | 54.4±5.4abcd | 6.6±0.7bcd | 15.3±0.9abc | 3.2±1.1ab | ||
| T4 | 50.3±5.7def | 7.0±0.7abc | 14.9±1.6bc | 3.8±1.2ab | ||
협수, 100립중, 수량을 조사하여 고온에 따른 수량 특성을 확인하였다(Table 3). 2021년 협수 16.7 ± 6.9개와 수량 55.6 ± 2.8 g으로 전생육기 T1처리에서 가장 높았고 온도 범위는 (최저온도 22.2°C; 평균온도 26.9°C; 최고온도 33.9°C)였다. 최저 수량을 나타낸 개화기 T4 처리에서 협수 4.3 ± 1.6개와 수량 6.8 ± 0.7 g였으며 온도 범위는 (최저온도 23.6°C; 평균온도 28.5°C; 최고온도 35.8°C)였다. 백립중은 영양생장기를 제외하고 전생육기, 개화기, 등숙기의 T4처리로 갈수록 감소하는 경향이 유의미하게 나타난다. 2022년 협수와 수량이 가장 높은 것은 등숙기 T2 처리에서 14.7 ± 4.4개와 57.5 ± 2.0 g는 온도 범위는 (최저온도 18.8°C; 평균온도 25.7°C; 최고온도 35.2°C)였다. 최저 백립중과 수량은 12.2 ± 1.0 g과 29.5 ± 3.4 g였으며 전생육기 T4 온도 범위는 (최저온도 22.9°C; 평균온도 28.8°C; 최고온도 36.9°C)였다. 또한 수량 최저범위가 개화기 T4 (최저온도 21.4°C; 평균온도 27.0°C; 최고온도 34.6°C)에서 나타났다. 수량과 협수는 온도에 영향을 받는데 최고, 최저 온도도 중요하지만 연차별 유사한 결과는 평균온도가 25.7 - 26.9°C 범위가 수량에 증진시키며 평균온도가 27.0 - 28.8°C이상이 되면 생육이 불량하고 수량성에 영향을 미쳐 수량이 낮아지는 경향을 보였다. 특히 ‘아라리’는 연차별 생육과 수량이 다른 품종보다 편차가 있는 것으로 보였으나(Chun et al., 2017; Oh et al., 2022) 다른 품종에 비해 수확량이 높은 편이여서 현재 팥 재배 품종 중 대다수를 차지하고 있다. 따라서 팥은 생육 적온 온도가 기존 보고된 온도(RDA, 2018)보다 상승한 것을 알 수 있었다. 특히 영양생장기에 최고 40.3°C까지 온도에서도 생육이 양호하였지만 개화기에는 평균 27°C 이상 되면 생육이 불량해지고 수량에 영향이 미치는 것을 이 실험을 통해서 알 수 있었다. 고온이 콩영양생장기의 생장 촉진 긍정적인 효과를 준다는 결과(Kantolic et al., 2013)와 콩 생식생장기 35°C이상의 고온에서 수량이 27%가 감소한 결과(Gibson & Mullen, 1996; Puteh et al., 2013)와 유사하게 팥에서도 고온은 영양생장기 보다 개화기에서 큰 영향을 미쳤고, 협수와 수량이 감소한 결과를 보였다(Table 3).
Table 3.
Yield components of “Arari” grown to high temperatures in 2021 and 2022.
| Year | Treatmentz | No. of pods | 100-seed weight (g) | Yield (g) | |
| 2021 | Con | 10.9±4.0bcy | 14.7±0.5abc | 41.1±0.9b | |
| WGP | T1 | 16.7±6.9a | 11.4±4.1de | 55.6±2.8a | |
| T2 | 12.0±4.2b | 10.7±3.0e | 39.8±4.5b | ||
| T3 | 10.5±2.7bcd | 13.9±0.5bcd | 29.6±3.8cdef | ||
| T4 | 8.3±4.7cdef | 12.1±1.1cde | 22.0±5.1f | ||
| VS | T1 | 8.6±3.6cdef | 14.7±0.8abc | 30.9±6.6cde | |
| T2 | 4.9±2.0gh | 17.4±1.5a | 21.8±2.0f | ||
| T3 | 6.6±3.1fgh | 16.8±0.7ab | 27.8±4.0cdef | ||
| T4 | 8.0±4.2def | 14.9±0.5abc | 25.3±7.0def | ||
| AS | T1 | 8.8±3.4cdef | 12.5±0.9cde | 29.8±4.7cdef | |
| T2 | 7.7±2.6def | 13.9±0.4bcd | 22.2±1.6f | ||
| T3 | 4.7±4.1gh | 12.4±0.9cde | 9.8±2.4g | ||
| T4 | 4.3±1.6h | 11.3±1.9de | 6.8±0.7g | ||
| MS | T1 | 10.4±2.7bcd | 14.0±0.3bcd | 36.1±2.6bc | |
| T2 | 9.7±4.1bcde | 14.1±0.9bcd | 33.8±6.6bc | ||
| T3 | 10.1±4.2bcde | 12.5±1.0cde | 33.3±5.6bcd | ||
| T4 | 7.2±3.2efg | 11.6±1.7de | 24.1±6.1ef | ||
| 2022 | Con | 12.8±5.2abc | 17.4±1.0ab | 53.7±9.9ab | |
| WGP | T1 | 11.9±4.6abcd | 16.1±0.8abcde | 46.6±5.9abcde | |
| T2 | 11.9±4.0abcd | 14.9±0.3e | 36.7±1.7def | ||
| T3 | 11.2±4.8abcd | 13.1±0.6ef | 30.8±2.6f | ||
| T4 | 11.7±5.9abcd | 12.2±1.0g | 29.5±3.4f | ||
| VS | T1 | 8.6±4.9d | 17.7±0.9ab | 33.7±2.0ef | |
| T2 | 9.6±4.2cd | 17.8±1.7a | 39.3±5.1cdef | ||
| T3 | 10.7±5.0bcd | 17.6±0.5ab | 46.1±1.4abcde | ||
| T4 | 11.4±6.0abcd | 17.8±1.1a | 41.0±14.4bcdef | ||
| AS | T1 | 11.7±6.5abcd | 17.3±0.7abc | 54.1±0.1ab | |
| T2 | 13.2±4.2abc | 15.7±0.7cde | 53.4±11.3ab | ||
| T3 | 11.7±4.0abcd | 15.4±0.1e | 46.7±11.9abcde | ||
| T4 | 10.3±5.4cd | 14.7±0.7e | 30.5±12.8f | ||
| MS | T1 | 12.3±4.5abcd | 17.1±0.7abcd | 51.7±3.4abc | |
| T2 | 14.7±4.4a | 16.1±0.8bcde | 57.5±2.0a | ||
| T3 | 14.6±4.4ab | 14.5±0.9ef | 54.5±4.5ab | ||
| T4 | 12.1±4.8abcd | 15.6±1.0cde | 48.3±3.7abcd | ||
팥의 일반성분 비교
2022년 생육시기별 고온 스트레스시 재배시 아라리의 종실 분석은 다음과 같다(Table 4). 대조구 아라리의 성분 함량은 수분은 각각10.2 ± 0.00%, 조회분은 3.9 ± 0.01%, 조단백질은 22.8 ± 0.13%, 총 식이섬유는 각 17.1 ± 1.12%, sucrose는 5.3 ± 0.11%로 보고된 연구에서 유사하거나 높은 범위였다(Chun et al., 2017; Oh et al., 2022; Sung et al., 2020). 단백질 양 25.7 ± 0.04%으로 전생육기 T4 (최저온도 22.9°C; 평균온도 28.8°C; 최고온도 36.9°C) 처리에서 가장 높은 값을 총 섬유질은 영양생장기 T4 (최저온도 25.2°C; 평균온도 30.5°C; 최고온도 37.9°C)에서 나타내는 것으로 보아 고온이 단백질과 식이섬유에 영향을 미치는 것으로 보인다. Sung et al. (2020)에 따르면 팥의 여러 품종은 일반적인 재배시 단백질 함량이 21.40 ± 0.22 - 23.30 ± 0.16% 였고 아라리는 23.03 ± 0.21%였지만 고온 스트레스시 가장 높은 값인 25.7 ± 0.04% 보였다. 이와 유사하게 결과는 콩 재배시 고온과 한발에서 스트레스가 높은 연도에 단백질이 1.1% 높게 나타났다(Shin et al., 2020). 또한 sucrose는 전생육기 T3, 개화기 T4, 등숙기 T2에서 높은 수치를 나타냈다. 식물의 재배시 재배환경, 생육상태와 종실 성숙과정에 따라 sucrose함량이 달라지고 고온일수록 유리당 중 sucrose의 함량 변화가 환경스트레스에 민감하게 좌우된다고 보고되었다(Krasensky & Jonak 2012).
Table 4.
Proximate composition results for “Arari” grown to high temperatures in 2022.
| Treatmentz | Moisture (%) | Ash (%) | Protein (%) | Total fiber (%) | Sucrose (mg/g) | |
| Con | 10.2±0.00hxy | 3.9±0.01b | 22.8±0.13k | 17.1±1.12a | 5.3±0.11ab | |
| WGP | T1 | 9.6±0.02k | 4.0±0.02a | 23.7±0.02d | 14.8±0.80bcd | 5.8±0.01ab |
| T2 | 9.4±0.04l | 3.9±0.02b | 24.2±0.01c | 14.1±0.33cdef | 5.1±1.42abc | |
| T3 | 9.1±0.09m | 3.8±0.02bc | 25.1±0.02b | 14.5±0.63bcde | 6.2±0.14a | |
| T4 | 8.9±0.07n | 3.8±0.02d | 25.7±0.04a | 15.3±0.27bc | 5.6±1.20ab | |
| VS | T1 | 11.0±0.02e | 4.0±0.04a | 23.0±0.10ij | 14.0±0.41def | 3.8±0.81c |
| T2 | 11.6±0.05c | 3.9±0.02b | 23.0±0.01j | 14.9±0.62bcd | 5.6±0.32ab | |
| T3 | 12.8±0.01a | 3.7±0.03ef | 23.4±0.07f | 15.0±0.58bcd | 5.3±1.25ab | |
| T4 | 12.7±0.00b | 3.7±0.02ef | 23.1±0.02hi | 17.4±0.86a | 5.9±0.19ab | |
| AS | T1 | 10.8±0.00g | 3.7±0.03ef | 22.8±0.06k | 13.2±0.72f | 4.5±0.38bc |
| T2 | 10.0±0.00i | 3.8±0.01cd | 23.1±0.03h | 13.5±0.41ef | 4.5±0.11bc | |
| T3 | 10.2±0.00h | 3.8±0.03b | 23.3±0.05g | 14.1±0.47def | 5.7±1.24ab | |
| T4 | 10.9±0.00f | 3.8±0.01de | 23.4±0.03f | 14.3±0.61cdef | 6.1±1.28a | |
| MS | T1 | 11.1±0.00d | 3.7±0.01f | 23.6±0.07de | 14.3±0.71cdef | 5.2±0.14abc |
| T2 | 10.2±0.00h | 3.7±0.01ef | 23.0±0.03j | 15.5±0.08b | 6.3±0.54a | |
| T3 | 10.0±0.00i | 3.7±0.03f | 23.1±0.06hi | 15.0±0.40bcd | 5.4±0.49ab | |
| T4 | 9.8±0.10j | 3.7±0.02f | 23.6±0.07e | 16.8±0.53a | 5.7±0.50ab | |
팥의 에탄올 추출물의 항산화성분의 함량
팥의 polyphenolic compounds는 항암과 항균의 기능을 하는 항산화 물질로 알려져 있으며 protocatechuic acid, coumaric acid 등 phenolic acid와 kaempferol, quercetin, myricetin등 flavonoid aglycone 또는 glycosides, proanthocyanidins 등이 포함되어 있다(Amarowicz et al., 2008; Song et al., 2014). 따라서 총 폴리페놀과 플라보노이드 함량을 측정하기 위해 에탄올로 추출하여 분석한 결과(Fig. 4) 생육시기와 고온에 따라 항산화성분이 유의적인 차이를 보였다. 대조구에서 총폴리페놀 및 총플라보노이드 함량은 132.1 ± 1.76, 130.1 ± 6.9 mg·100g-1로 기존 연구에서보다 높은 수치를 보였다(Chun et al., 2017; Sung et al., 2020). 전생육기와 영양생장기의 고온 스트레스시 총폴리페놀과 총플라보노이드 함량이 증가하여 논 토양 재배로 수분스트레스 받았던 연구결과(Chun et al., 2017)보다 높은 수치인 256.9 ± 11.08와 272.7 ± 15.7 mg·100g-1 나타났다. 토마토는 고온 스트레스를 받게 되면 총폴리페놀 및 총플라보노이드 함량은 급증하고(Alhaithloul et al., 2021), 팥 연차별 노지 기상 조건의 일조 시간이 적었던 해에서 항산화 성분과 활성이 높았던 것을 확인하였다(Oh et al., 2022). 따라서 팥은 고온 스트레스하에서도 총폴리페놀 및 총플라보노이드 함량이 유의미하게 증가하여 생육 과정 중 고온 및 습해 스트레스 혹은 적은 일조시간의 영향을 받게 되면 식물체에 항산화 성분이 증가함을 알 수 있었다.
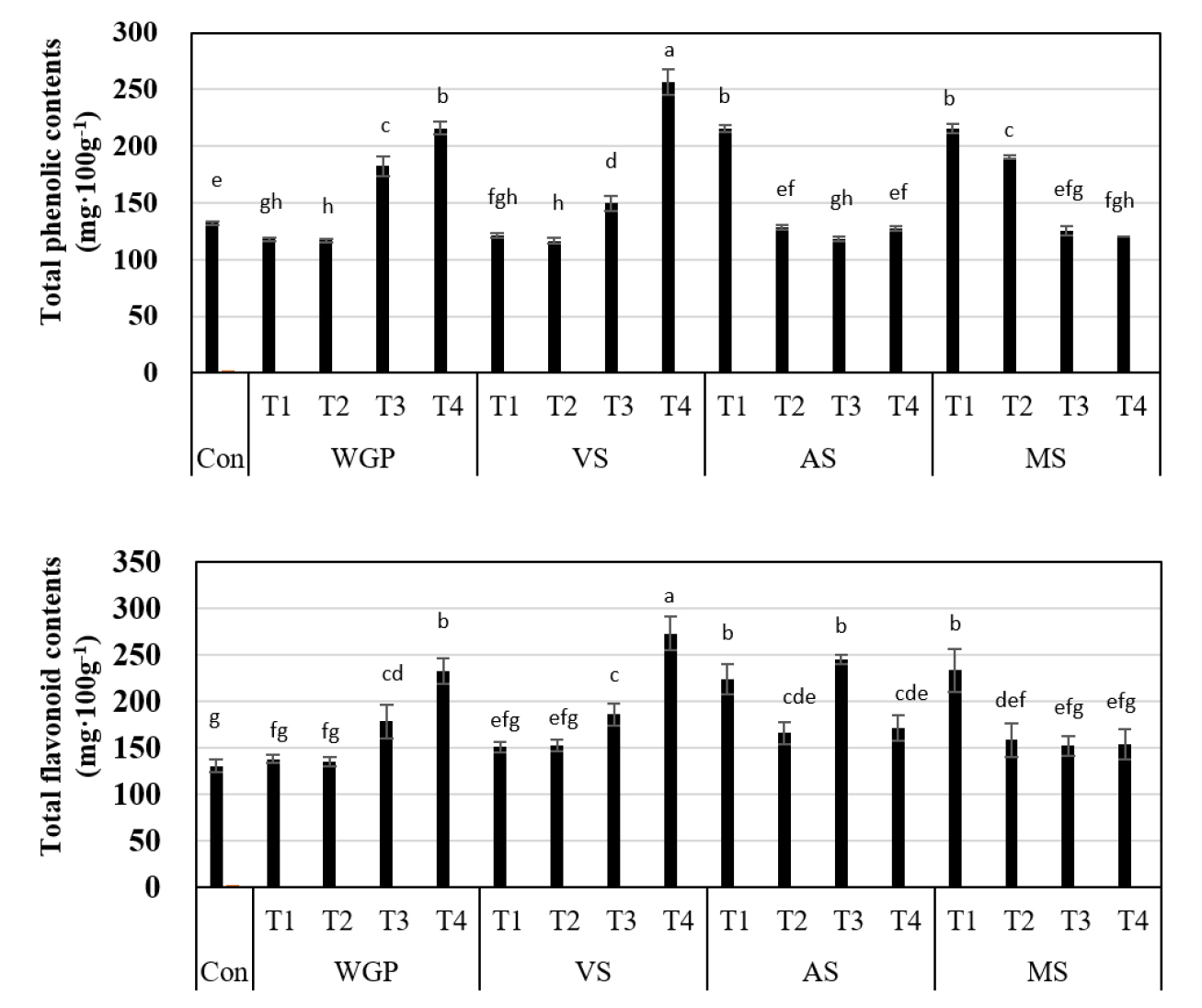
Fig. 4.
Comparison of total phenolic contents and total flavonoid contents of “Arari” in 2022. Error bars indicate standard deviations and differences were tested with one-way ANOVA, as indicated with letters, using Duncan’s multiple tests (p < 0.05). T1: control + 1°C, T2: control + 2°C, T3: control + 3°C, T4: control + 4°C.
생육시기에 따른 고온 스트레스시 화분 형태와 활력 비교
전생육기에서 화분 형태와 임성 확인은 주사전자현미경(Scanning electronic microscopy; SEM)과 Alexander 염색약으로 확인하였다(Peterson et al., 2010). 팥 화분의 형태 용어는 다음의 논문에서 참고하였다(Kim et al., 2020; Koti et al., 2005; Umdale et al., 2017). 극축면에서는 아원형과 원형의 형태이고 적도면에서는 아단구형 약장구형(prolate)의 과립상 돌기를 지닌 3공구형(tricolporate)의 발아구를 갖고 있다. 화분표면무늬는 그물모양(reticulate-heterobrochate) 특징 가지고 있고 lumina 형태는 polygonal이고 muri는 narrow-straight를 가지고 있다(Fig. 5A). 기존의 연구에서 팥의 화분 특징이 ‘아라리’에서도 유사하게 나타나고 크기와 특성이 대조구와 고온시 변화하는 것을 확인하였다(Umdale et al., 2017). 비정상적인 화분은 비균일한 모양이거나 찌그러져 있거나 덜 발달하여 크기가 작은 형태를 보이고 있다(Fig. 5B and C). SEM사진의 화분 형태(Fig. 5D, E and F)와 Alexander 염색(Peterson et al., 2010)으로 비정상 화분(Fig. 5G, H and I)을 구분한 결과 대조구와 전생육기 T1 (최저온도 22.1°C; 평균온도 27.0°C; 최고온도 33.9°C)에서는 통계적으로 차이가 없었지만 T4 (최저온도 22.9°C; 평균온도 28.8°C; 최고온도 36.9°C)에서는 급격히 정상 화분(45.6%과 40.8%)이 감소했다(Fig. 5J and K). Faba bean에서 32°C 고온처리시 화분의 50%의 발아가 억제되어 수량이 감소한 연구결과(Bishop et al., 2016)와 콩의 생식생장시기의 고온 스트레스가 수량에 영양을 미친다(Puteh et al., 2013)는 결과와 유사하게, 고온이 팥의 화분 임성을 감소시키고, 결과적으로 수량 감소에 영향을 미치는 것으로 판단된다.
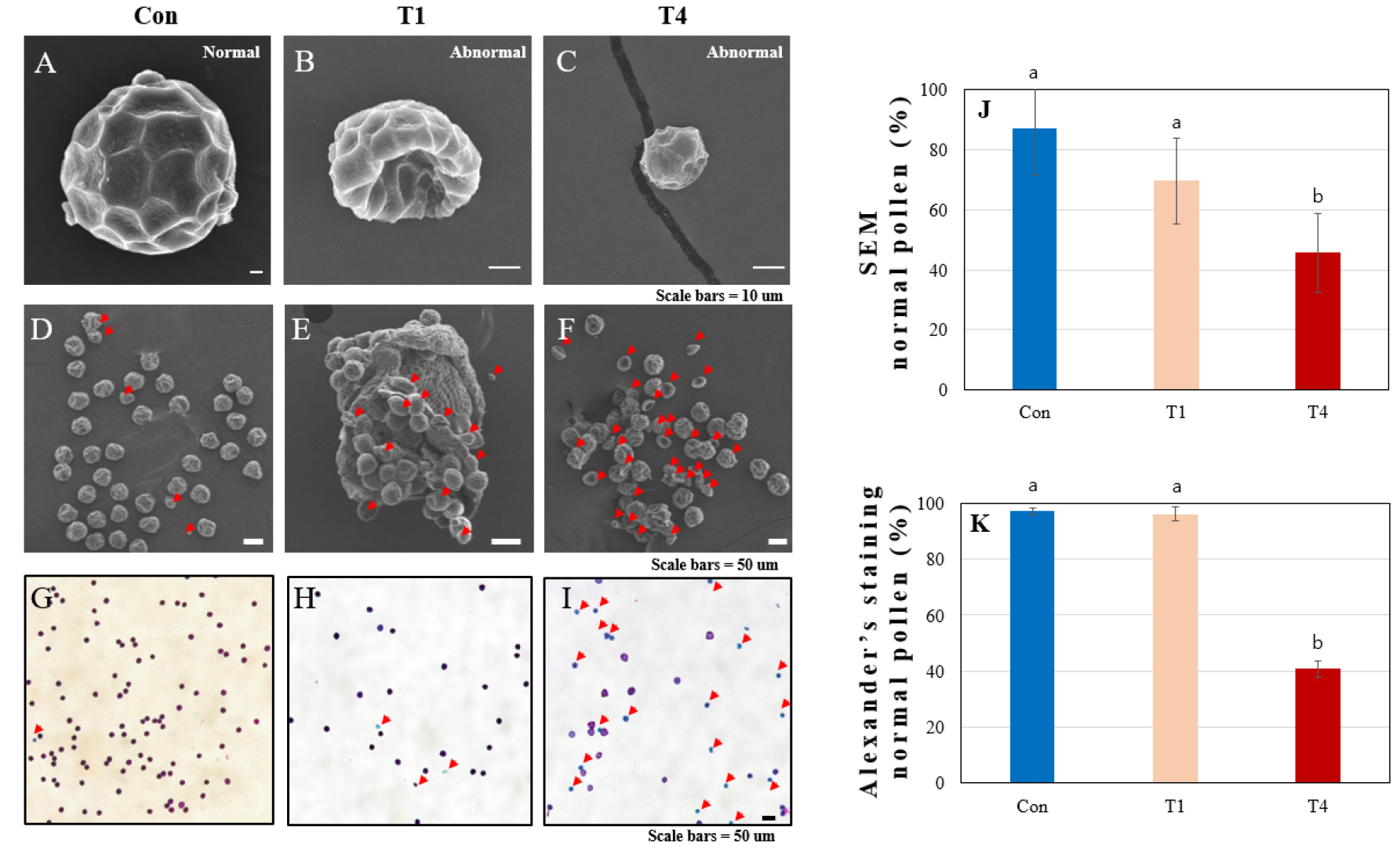
Fig. 5.
The pollen morphology and pollen viability of V. angularis cv. “Arari” at high temperatures in 2022 (A - F: Scanning electron microscopy (SEM) of pollen grains by high-temperature treatment control, T1 and T4; G-I: Alexander’s staining of pollen grains by high temperature treatment control, T1 and T4; J and K: The proportion of normal pollen by scanning electron micrographs (SEM) and Alexander’s staining (Con: rain shield greenhouse, T1: control + 1°C, T4: control + 4°C). The red arrows indicate abnormal pollen. Error bars indicate standard deviations, and differences were tested with one-way ANOVA, as indicated with letters, using Duncan’s multiple tests (p < 0.05).
적 요
본 연구는 최근 빈번한 고온 노지 재배 피해가 빈번함에 따라 팥 대표 품종 ‘아라리’의 2021과 2022년 고온구배온실에서 수행된 고온 반응 실험이다. 적정 온도 보다 높은 온도 범위에서 팥의 노지 재배시 고온 연구로 국내 품종 ‘아라리’의 생육 반응, 수량, 화분 활력, 종실 성분 등을 최초로 비교분석 하였다.
1. 세부적인 기상 조건은 2021과 2022년의 최저, 평균, 최고기온 연차간 차이는 2022년 보다 2021년이 전생육기는 0 - 1.0°C로 차이가 적었고 영양생장기 0 - 3.7°C, 개화기는 0.4 - 2.4°C로 높았다. 2021년 보다 2022년에서 등숙기는 최저, 평균 온도는 낮았지만 최고온도는 1.7 - 3.9°C로 높아 연차별 온도에 따라 생육특성이 다르게 나타났다.
2. 연차별 생육기간을 비교해 보면 개화기는 평균 기온차에 의해 2021년 13일, 2022년 27일로 차이가 있었으며 등숙기는 2021년 30일, 2022년 19일로 차이가 있었다. 영양생장기에는 최고 온도가 40.3서도 생육이 양호하였지만 개화기 고온 처리는 평균온도가 27.0°C 이상이 되면 생육이 불량하여 수량이 낮아지는 경향을 보였다. 2021년에 비해 2022년 등숙기 온도가 최저 온도 1.6 - 1.9°C 낮고 최고 온도 1.7 - 3.9°C로 높고 기간이 늘어나니 팥의 수량성이 높았다.
3. 전생육기와 영양생장기의 고온 스트레스시 총폴리페놀과 총플라보노이드 함량이 증가하였고 화분 형태와 활력은 대조구와 전생육기T1에서는 통계적으로 차이가 없었지만 T4 (최저온도 22.9°C; 평균온도 28.8°C; 최고온도 36.9°C)에서는 비정상 화분 형태가 45.62%, 급격히 화분 활력이 40.75%로 떨어졌다.
4. 고온 스트레스시 수량은 2021년 개화기 T4 (최저온도 23.6°C; 평균온도 28.5°C; 최고온도 35.8°C)에 2022년은 수량과 화분 임성은 전생육기 T4 (최저온도 22.9°C; 평균온도 28.8°C; 최고온도 36.9°C)에 가장 낮았으며 반면 총폴리페놀과 총플라보노이드는 영양생장기 T4 고온 스트레스가 높은 시기일 때 가장 높았다.
