

서 론
재료 및 방법
재배환경 설정
시험재료 및 재배관리
조사방법
통계처리
결과 및 고찰
온도구배하우스 내 감자 재배환경
온도상승에 따른 감자의 생육변화 분석
온도상승에 따른 감자의 수량변화
온도상승에 따른 감자의 생육 스트레스 분석
고온스트레스 시 생육변화 분석을 위한 최적 분석방법 탐색
적 요
서 론
최근 기후변화 및 기상이변으로 고온, 한발, 습해와 같은 기상스트레스의 발생 빈도 및 강도가 지속적으로 증가하고 있으며, 2013년 3월(7℃)과 4월(11℃)의 평균기온에 비해 2023년에는 각각 약 3℃ 상승한 10.2℃와 14.2℃를 기록한 것으로 나타났다(KMA, 2024; KOSIS, 2024).
고온 환경은 봄 감자의 초기 생장을 촉진하지만, 후기 생육과 수량 감소의 원인이 된다(Lee et al., 2020). 감자의 생육 적온은 14℃~22℃로 알려져 있으며 적온 조건에서는 괴경형성이 촉진되고 줄기발달이 억제되는 반면, 고온에서는 잎의 건물량이 증가하고, 괴경 건물량과 크기가 감소하여 감자의 품질과 생산성을 심각하게 저하시킬 수 있다(Singh et al., 2020). 이러한 현상은 감자의 생산성과 품질을 저하시키는 주요 요인으로 작용하고 있으며 실제 감자의 생산량은 2023년 기준 10a당 2,436 kg으로, 지난 10년간 2,722 kg(2013년 기준)에서 꾸준히 감소한 것으로 나타났다(KOSIS, 2024).
최근 디지털 기술의 발달로 영상과 센싱 장비를 이용하여 고온 조건과 같은 기상재해를 받은 작물의 생육 이미지의 시계열 자료와 작물 센싱 데이터 등의 빅데이터 생산이 가능해졌으며, 인공지능 분석 기술을 적용 시 작물의 스트레스를 조기에 판별, 진단할 수 있다(Sun et al., 2022). 작물의 생육 스트레스를 분광 및 센서로 측정 시 NDVI (Normalized Difference Vegetation Index), PRI (Photochemical Reflectance Index), CWSI (Crop Water Stress Index)와 같은 생육지수를 산출할 수 있다. NDVI는 식물 전반적인 생육 상태를 모니터링하여 건강 상태와 활력을 평가하는데 도움을 줄 수 있으며, PRI는 광합성 효율을 측정하는 지표로, 고온 스트레스로 인한 광합성 활동 저하 정도를 평가할 수 있다(Chen et al., 2020; Lazarević et al., 2022). 또한 CWSI는 증산작용과 같은 식물의 생리적 반응을 평가하여 고온 스트레스로 인한 수분부족에 대한 반응을 이해할 수 있다(Rud et al., 2014).
감자는 넓은 재배면적과 높은 생산량에도 불구하고 상대적으로 디지털 기술을 접목한 생육진단에 대한 연구가 미흡한 실정이다. 최근 기상스트레스가 가중되고 있는 상황에서 스트레스에 따른 감자 피해를 효율적으로 진단·분석하여 피해를 조기에 진단하고 대응할 수 있는 기술의 개발이 필요하다. 이에 본 연구는 고온 조건에서 나타나는 감자 품종별 생육변화와 생산성 변화를 분석하고, 최신 디지털 영상과 센싱기술을 접목하여 기본 생육조사 및 디지털 센싱기술을 통한 수확 예측의 가능성을 평가하고자 한다.
재료 및 방법
재배환경 설정
다양한 온도 조건에서 생육 반응을 관찰하기 위해 전라북도 완주군 이서면 국립식량과학원에 위치한 온도구배하우스 시설(Fig. 1)을 이용하여, 시험기간 동안 터널 입구에서 온∙풍구 방향으로, 온도가 점차적으로 상승하도록 하여 온도를 제어하였다. 온도와 습도는 6 m 간격으로 설치되어 있는 4개의 온습도 센서를 이용하여 측정되었으며(Fig. 2), 온∙습도 및 일사량은 온도구배하우스 내 환경 조건을 실시간으로 모니터링하기 위해 자동으로 측정·기록되는 제어실에서 추출한 데이터를 이용하였다.
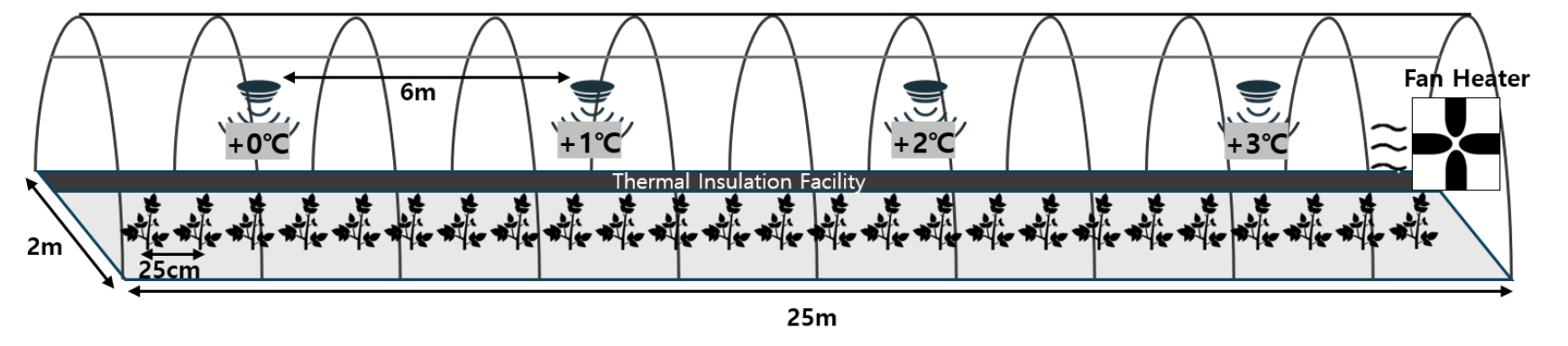
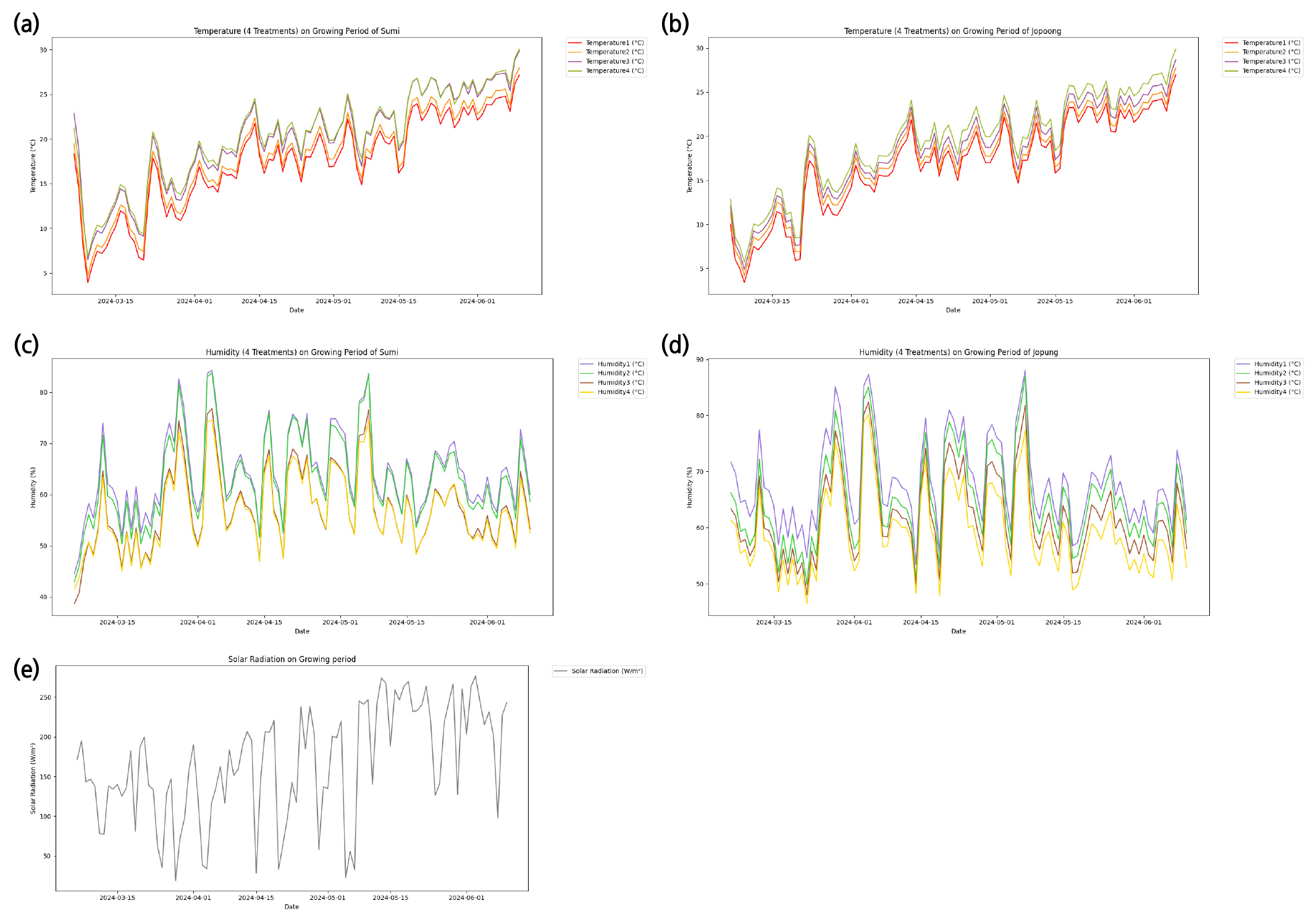
Fig. 2.
Daily variation of air temperature in Sumi (a), air temperature in Jopung (b), Relative Humidity in Sumi (c), Relative Humidity in Jopung (d), Solar Radiation (e) for growing potato (Solanum tuberosum L.) cultivation during the experimental period in the Thermal Gradient House (Mar.-Jun. 2024).
시험재료 및 재배관리
본 실험에 사용한 감자(Solanum tuberosum L.)는 조생종인 ‘수미(Sumi)’와 ‘조풍(Jopung)’으로, 농촌진흥청 고령지 농업연구소에서 씨감자를 분양받았다. 파종할 감자는 280 g 이상의 건전하며, 싹 세력이 균일한 씨감자를 선별한 후 절단 및 소독하였다.
싹 틔운 감자는 3월 초순에 두 품종을 각각 3동씩, 총 6동에 정식하였다. 각 온실 내 골은 한 골로 하여, 한 골에 총 100개체씩 재식 간격은 25 cm로 하였다. 시비방법은 전량기비하였고, 농촌진흥청 표준시비법을 따랐다(RDA, 2020). 조사구 지점은 각 동에서 6지점씩 일정한 간격으로 개체를 선별하였다. 각 개체별 온도차이는 온풍구로부터 각 온습도 센서 및 조사구 지점까지의 거리를 이용하여, 외기온도 대비 상승한 온도 차이(℃)를 계산하였다.
조사방법
다양한 온도 조건에서의 작물의 생육을 관측하기 위해 주요 밭작물 재배기술 및 시험연구 조사기준 필드북(NICS, 2021)을 기준으로 초장, 엽수를 측정하였으며, 해당 기준에 따라 수량을 무게 기준별로 분류하여 상서수량을 측정하였다. 엽색은 SPAD-502 Plus (Konica Minolta Optics, Japan)을 이용하여 Li et al. (2012)의 방법에 따라 상위 4번째 엽을 측정하였다.
고온에 따른 작물의 스트레스를 확인하기 위해 NDVI, PRI, CWSI 등의 식생지수를 활용하였다. NDVI는 하향 센서인 S2-412 NDVI sensor (Apogee instruments, USA)와 상향센서인 S2-411 NDVI sensor (Apogee instruments, USA)를 로고 센서와 연결하여 오전 10시~12시에 이동하면서 측정하였다.
PRI는 하향 센서 S2-422 SRS-Pr PRI sensor (Apogee instruments, USA)와 상향 센서 S2-421 SRS-Pi PRI sensor (Apogee instruments, USA)를 이용하여 광합성이 활발히 일어나는 오전 10시~12시에 이동하면서 측정하였다.
CWSI를 측정하기 위해서는 SI-431 IR sensor (Apogee instruments, USA)를 이용하였다. IR senor는 한 동에서 6지점씩 설치하여 생육을 관찰하였다. 30분 간격으로 측정하였으며, 온도의 영향을 가장 많이 받는 오후 1시~2시에 측정된 데이터를 이용하였다. CWSI 지수를 추출하기 위해 엽온과 대기온도의 차(Tc-Ta), 포화수증기압차(Vapor Pressure Deficit, VPD)를 사용하였다. 엽온은 각 처리구마다 지표면으로부터 100 cm 높이, 직하(0°)로 IR센서(SI-431, Apogee instuments)를 설치하여 30분 간격으로 측정하였으며, 대기온도와 상대습도는 온도구배하우스 시설의 제어실에서 자동측정되는 데이터를 이용하였다. CWSI 값의 산출은 경험적 방법(Jackson et al., 1981)을 이용하였다.
=Intercept + Slope (VPD),
=Intercept + Slope (+ Intercept))
통계처리
본 연구의 모든 처리는 3반복 이상 평균으로 계산되었으며, 실험데이터는 Python (Version 3.7.16)을 이용하여 일원분산분석(ANOVA)를 실시하였고, 1% 및 5% 수준에서 통계학적 유의성을 검정하였다.
결과 및 고찰
온도구배하우스 내 감자 재배환경
전지구 기후변화 전망보고서(NIMS, 2014)에 따르면 2014년 대비 2021~2040년에 1.6~1.9℃, 2081~2100년에 2.7~7.6℃로 온도가 상승하게 된다. 생육 후기에 최고기온이 30℃ 이상인 일수가 증가하게 되면 높은 호흡량으로 인해 감자의 괴경 형성 및 비대를 저해할 수 있기 때문에 지구온난화로 인한 고온은 괴경 수량에 부정적인 영향을 미칠 수 있다(Kim & Lee, 2016).
온도구배하우스 내 감자 재배기간 동안의 일일적산량은 기상이 좋지 않은 날을 제외하였을 때 3월은 3419.19 Wh/m2, 4월은 4062.22 Wh/m2, 5월은 5225.12 Wh/m2, 6월은 5083.8 Wh/m2였다(Fig. 2e). 재배기간 동안의 수미의 온도구배하우스 입구의 일평균기온은 17.7℃, 후미의 일평균기온은 22.0℃였으며(Fig. 2a), 조풍은 입구 일평균기온이 18.3℃, 후미의 일평균기온은 21.2℃였다(Fig. 2b). 입구로부터 후미로 갈수록 온도가 상승하게 하였다.
온도상승에 따른 감자의 생육변화 분석
감자는 고온스트레스를 받게 되면 생육이 저하되고, 생육 후기 노화가 빨라진다(Ramírez et al., 2014). 고온의 영향을 받은 감자의 생육변화를 예측하기 위해 온도구배하우스 내에서 감자를 재배하여 생육과 관련된 지표인 감자의 초장, 엽수 및 잎 엽록소 함량변화를 조사하였다(Figs. 3, 4, 5).
먼저 감자 재배기간 동안 온도 상승에 따른 초장 변화와 생육기간 동안 평균 초장 변화를 분석한 결과, 두 품종 모두 파종 후 62일까지 생육이 진전됨에 따라 초장이 증가하였다(Fig. 3). 수미는 생육기간 동안 평균 초장은 온도구배 하우스 입구 기준 1.6℃ 상승했을 때 가장 높은 값을 보인 것으로 보아 수미는 1.6℃ 이상 상승하게 되면 고온스트레스로 생육이 감소한다는 것을 알 수 있었다(Fig. 3a, c). 조풍의 경우 파종 후 49일까지 초장이 온도상승에 선형적으로 증가하였으며, 생육기간 동안 가장 높은 처리인 3.3℃ 상승하는 경우 초장이 가장 컸다(Fig. 3b). 생육기간 동안 평균 초장값 또한 온도가 높아질수록 증가하는 것으로 나타났다(Fig. 3d).
감자 재배기간 동안 온도 상승이 엽수에 미치는 영향과, 재배기간 동안의 평균 엽수를 분석한 결과, 수미는 생육이 진전됨에 따라 엽수가 증가하다가 파종 후 49일에 가장 높은 값을 보였으며(Fig. 4a), 재배기간 동안 평균 엽수는 1.6℃ 상승했을 때 가장 높았다(Fig. 4c). 조풍이 엽수가 최대에 이르는 시기인 62일까지 엽수가 온도와 양의 상관관계로 증가하는 모습을 보였으며, 재배기간 동안 엽수 평균은 3.3℃ 상승했을 경우 가장 컸다(Fig. 4b, d).
지상부의 생육을 보았을 때 수미가 전 생육기간 동안 지상부 생육(Fig. 3a, c)이 고온스트레스를 받지 않고 안정적으로 자랄 수 있는 온도는 1.6℃ 이하에서 상승하는 경우였다. 조풍(Fig. 3b, d)의 경우 고온이 지상부 생육과 긍정적으로 작용하는 것을 알 수 있었다.
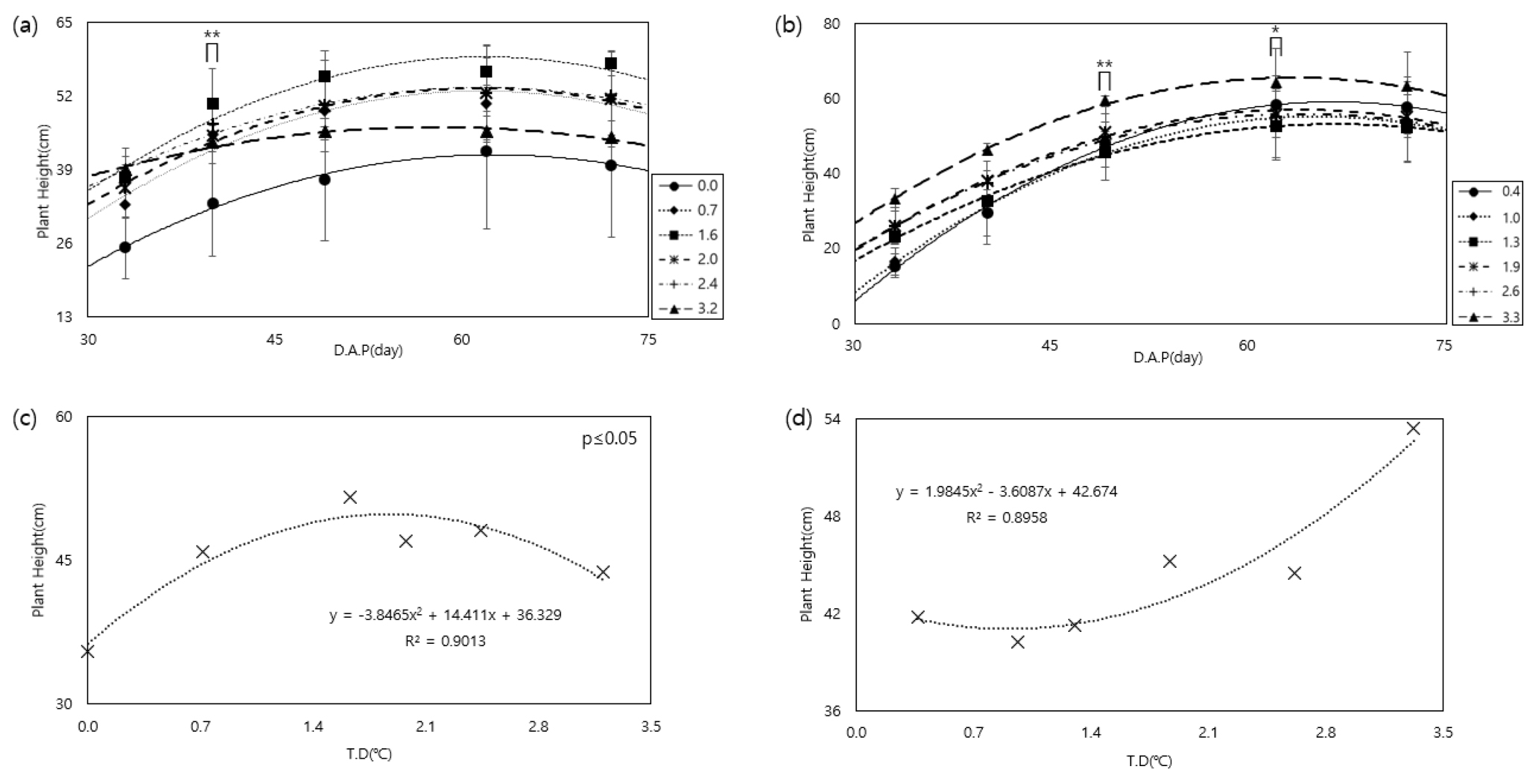
Fig. 3.
Changes in plant height according to temperature increase during two potato varieties cultivation in a temperature gradient house ((a): Sumi, (b): Jopung) and comparison of average in plant height during cultivation according to temperature treatment((c): Sumi, (d): Jopung). ∏ Significance level: 0.01*, 0.05**. * D.A.P (day) : Days After Planting. * T.D (°C): Temperature Difference compared to external conditions.
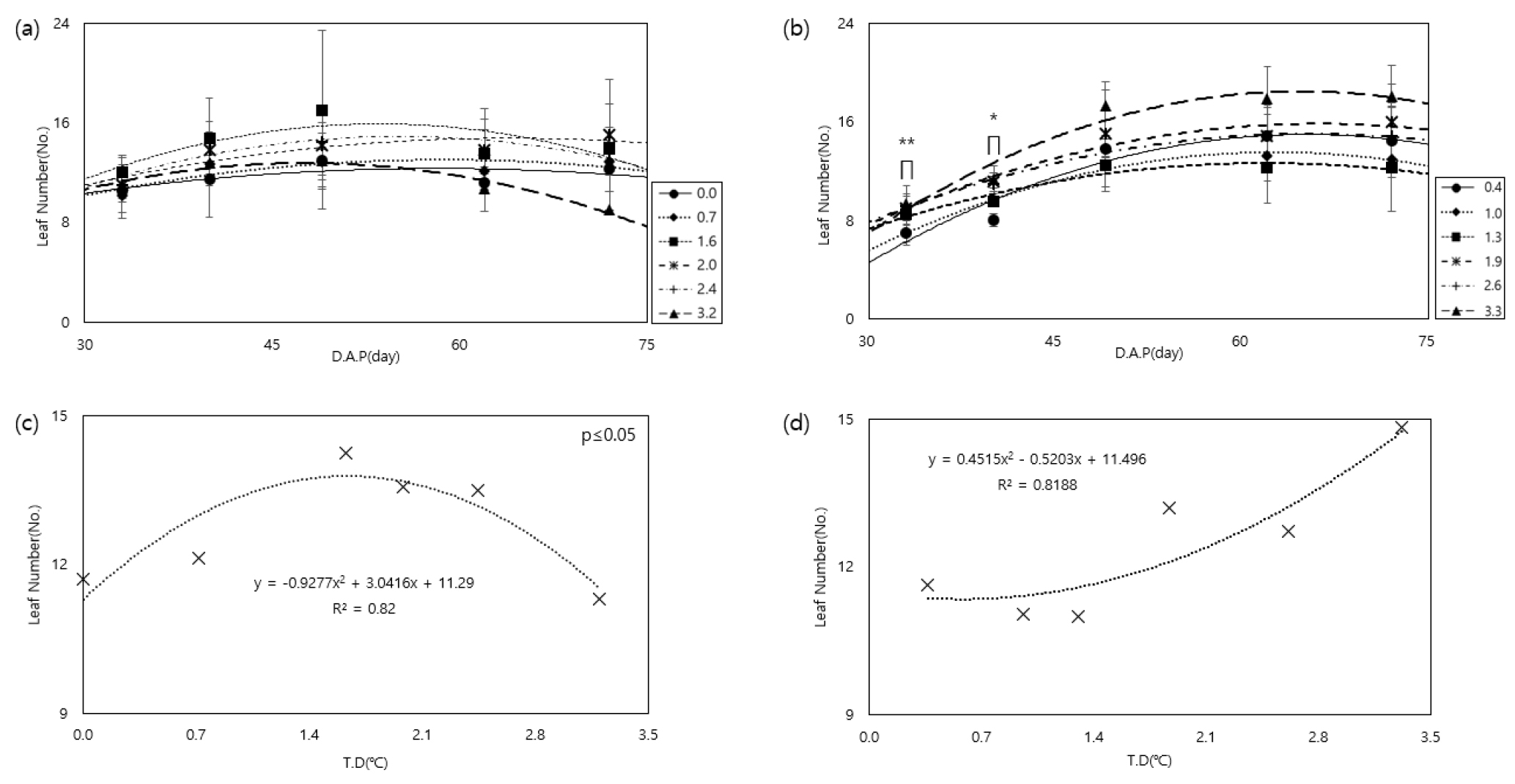
Fig. 4.
Changes in leaf age according to temperature increase during two potato varieties cultivation in a temperature gradient house ((a): Sumi, (b): Jopung) and comparison of average in leaf number during cultivation according to temperature treatment((c): Sumi, (d): Jopung). ∏ Significance level: 0.01*, 0.05**. * D.A.P (day): Days After Planting. * T.D (°C): Temperature Difference compared to external conditions.
잎이 고온에 노출될 경우, 잎을 대부분 구성하고 있는 엽록소가 열에 민감하기 때문에 엽록소가 분해되고, 엽록체 구조가 손상될 수 있다(Zahra, 2023). 이는 결국 광합성 활동을 저해하여 괴경 생산량을 저하시킬 수 있다. 온도상승이 SPAD값에 미치는 영향을 분석하기 위해 감자 재배기간 동안 SPAD값 변화를 조사하고, 재배기간 동안의 평균 SPAD 값을 비교하였다(Fig. 5). 두 품종 모두 괴경이 형성되는 시점인 파종 후 40일부터 SPAD 값이 감소하는 경향을 보였다(Fig. 5a, b). 수미는 재배기간 동안 평균 SPAD 값이 최대를 보였던 것은 1.6℃ 상승하는 처리였고 1.6℃ 이상 상승했을 때 감소하는 경향을 보였다(Fig. 5c). 조풍은 수미보다 SPAD값의 범위를 높은 수준으로 유지하였는데 0.4℃ 상승했을 때, 그 이상의 온도에서는 처리 간의 차이가 거의 없었다(Fig. 5d). 열 스트레스에 비교적 잘 견딜 수 있는 경우 초장과 엽록소가 증가한다고 한다(Tang, 2018). 조풍은 수미에 비해 지상부의 생육 정도가 크고, SPAD값의 범위가 더 높은 것으로 보아 온도에 대한 적응성이 있어 보이는 것으로 판단된다. 조풍이 수미에 비해 고온스트레스를 적게 받는 것으로 보이는 것은 봄·여름·가을·시설재배 등이 모두 가능한 적응성이 큰 품종이기 때문에 온도에 대한 적응성 또한 높은 것으로 추측된다(RDA, 2020).
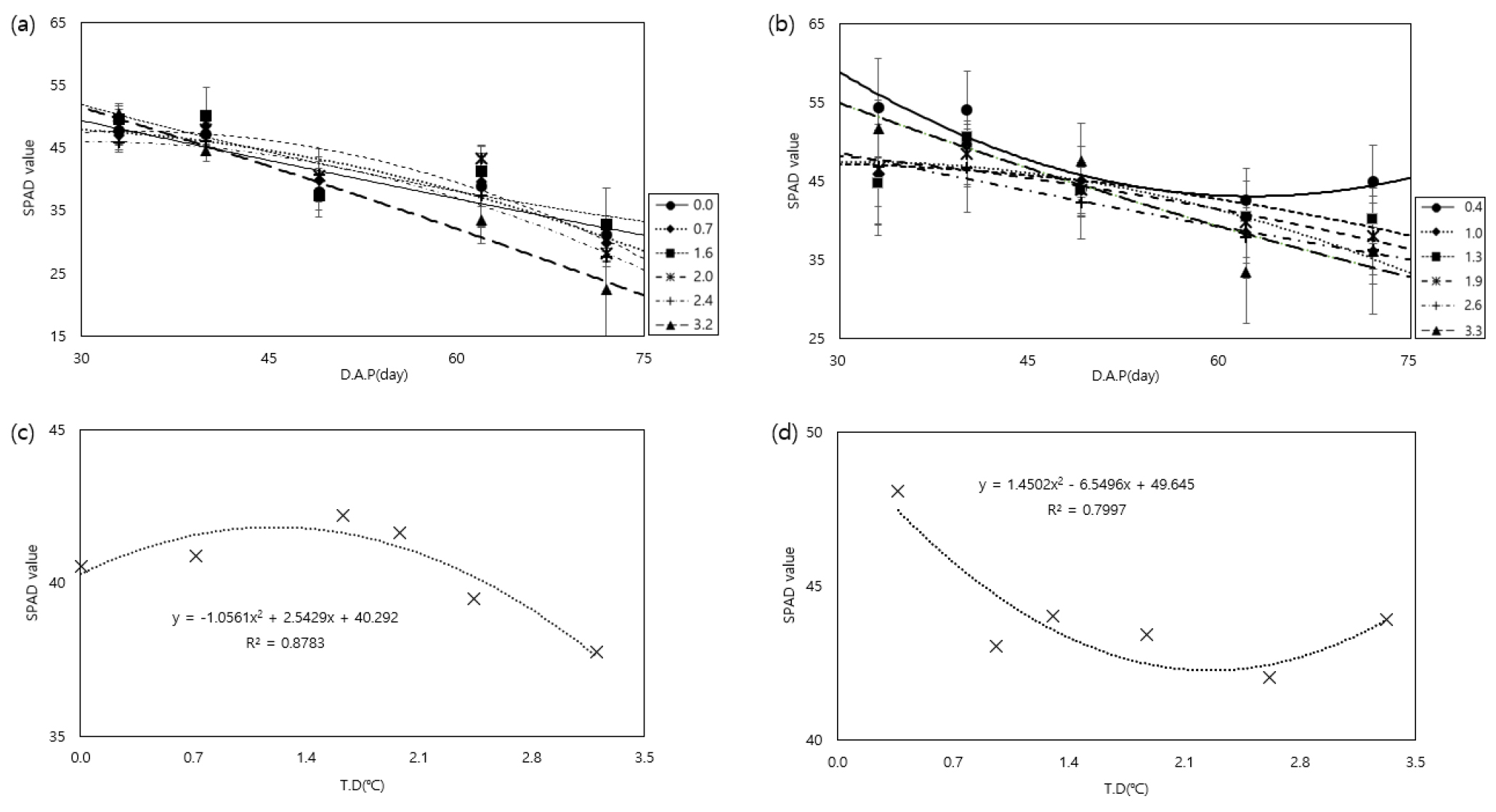
Fig. 5.
Changes in SPAD value according to temperature increase during two potato varieties cultivation in a temperature gradient house ((a): Sumi, (b): Jopung) and comparison of average in SPAD value during cultivation according to temperature treatment((c): Sumi, (d): Jopung). * D.A.P (day): Days After Planting. * T.D (°C): Temperature Difference compared to external conditions.
온도상승에 따른 감자의 수량변화
감자는 고온의 환경에서 스트레스를 받게 되면 수량이 감소하게 된다(Marinus & Bodlaender, 1975). 온도상승이 감자 두 품종의 상서수량에 미치는 영향을 조사하였다(Fig. 6). 수미에서는 온도가 상승할수록 수량이 증가하다가 일정 온도 이상에서는 감소하는 경향을 보였는데, 최대 수량을 보인 온도는 하우스 입구 기준 상승한 온도범위 0.0℃~3.0℃에서 2.4℃ 상승한 처리였다. 조풍의 경우 수미보다 수량 생산이 많은 편이었는데, 온도 상승에 따른 수량 변화는 없었다. 지상부의 생육(Figs. 3, 4)은 온도와 긍정적으로 반응했음에도 수량이 증가하지 않는 결과에 대해서는 생리적인 반응 및 이화학적 특성에 대한 추후 실험이 필요해 보인다. Lee et al. (2022)에 따르면, 감자 5품종에 대해 폭염 적응성 실험을 한 연구에서도 조풍이 수미를 포함한 다른 품종들에 비해 상서수량 감소가 낮았다고 한다.
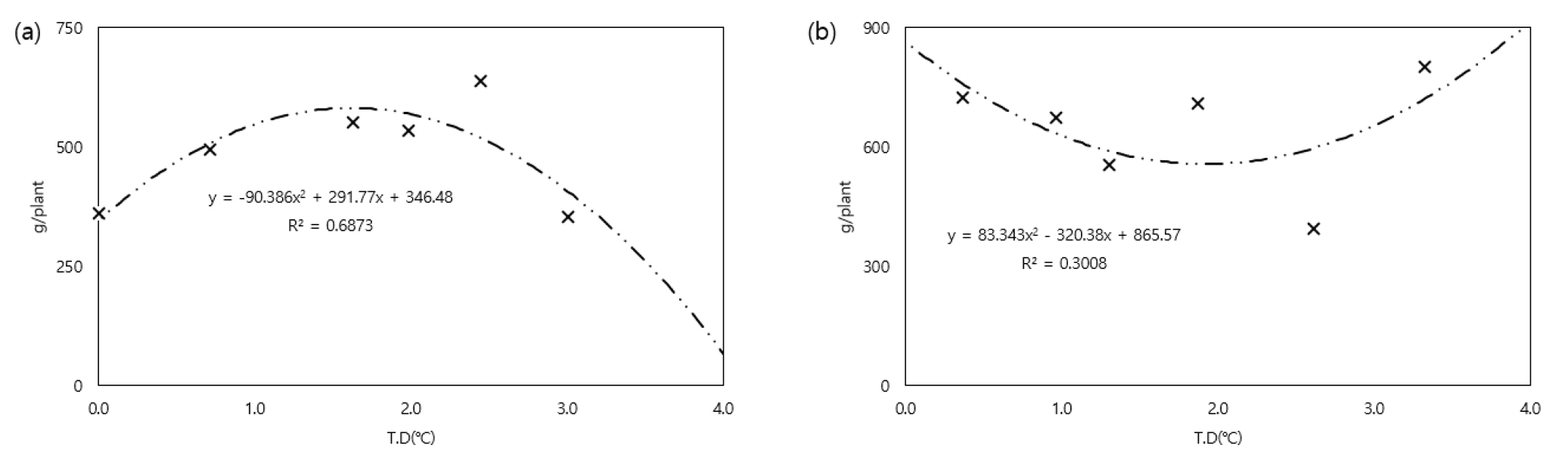
온도상승에 따른 감자의 생육 스트레스 분석
감자의 수량은 생육단계 중 괴경비대기의 생육정도에 영향을 가장 많이 받는다고 알려져 있는데, 이 시기는 지상부 생육의 최성기로 간주된다(Luo, 2020). 따라서 괴경 비대기이자 생육최성기의 온도 조건에 따른 생육스트레스 정도를 디지털 기술을 활용하여 진단하기 위해 파종 후 58일~72일 동안 센서로 측정한 각 생육지수의 평균 값들을 분석하였다(Fig. 7).
지상부의 생육이 최대인 시기 동안의 NDVI값의 평균 값을 조사한 결과, 두 품종 모두 온도에 따른 경향이 나타나지 않았다(Fig. 7a, b). NDVI 분석결과는 앞서 생육분석에서 측정한 SPAD값과 차이를 보였는데 이는 SPAD의 경우 광합성이 가장 활발히 일어나는 상위 엽 네 번째 잎의 한 지점을 측정하지만(Li et al., 2012), NDVI는 식생에서 반사되는 적색광과 근적외선 파장대의 값을 토대로 한 개체의 전반적인 생육 상태로 엽록소 농도를 추정하기 때문인 것으로 판단된다(Lazarević et al., 2022). NDVI와 SPAD 분석 결과, 3.2℃ 이하로 온도가 상승했을 때 엽록소 함량 변화는 센서를 기반으로 분석이 가능하지만 작물의 전체 표현형으로 고온에 의한 스트레스를 추정하는 것은 한계가 있는 것으로 판단하였다.
다양한 온도범위가 광합성 효율에 미치는 영향을 조사하기 위해 괴경비대기 동안 일사량이 좋았던 날의 오전 10~ 12시의 평균 PRI값을 확인하였다(Fig. 7c, d). 수미의 괴경비대기 동안 PRI 평균 값은 온도가 증가할수록 증가하다가 1.6℃ 이상 상승했을 때 감소하였다. 이는 생육과 수량에서 또한 유사한 경향을 보인 것으로 보였다(Fig. 3, 4, 5, 6). 조풍은 수미에 비해 PRI 값의 범위가 높은 수준으로 유지되는 것으로 보아 광합성이 상대적으로 활발해보였는데, 온도 상승에 따른 경향은 보이지 않는 것으로 보아 광합성 효율에 대한 스트레스는 거의 없는 것으로 판단된다.
작물이 고온스트레스를 받아 호흡이 왕성해지면, 증산활동이 활발해져 작물이 고온에 의한 2차 피해로 수분스트레스를 받을 수 있다(Ierna, 2023). 또한 괴경비대기는 생육단계 중 수분에 가장 민감한 시기이다(Van Loon, 1981). 고온이 CWSI값에 미치는 영향을 분석하기 위해 괴경비대기 동안 일사량이 좋았던 날에서 하루 중 가장 기온이 높은 오후 1시~2시의 평균 CWSI 값을 분석하였다(Fig. 7e, f). 수미는 모든 온도에서 평균 CWSI 값이 높은 수준을 유지하는 것으로 보아 수분스트레스를 받고 있지만, 온도 처리 간의 경향성은 나타나지 않았다. 조풍은 수미에 비해 낮은 CWSI값을 유지하였는데, 온도가 높아질수록 CWSI 값이 증가하는 경향을 보였다. 수미와 조풍의 각 상대습도를 확인한 결과 습도 차이가 거의 나지 않은 것으로 보아(Fig. 2), 습도차이로 인한 품종 간의 상이한 결과가 아닌 고온스트레스로 인한 활성화된 대사활동에 따른 2차 피해의 결과로 판단할 수 있다(Fig. 2).
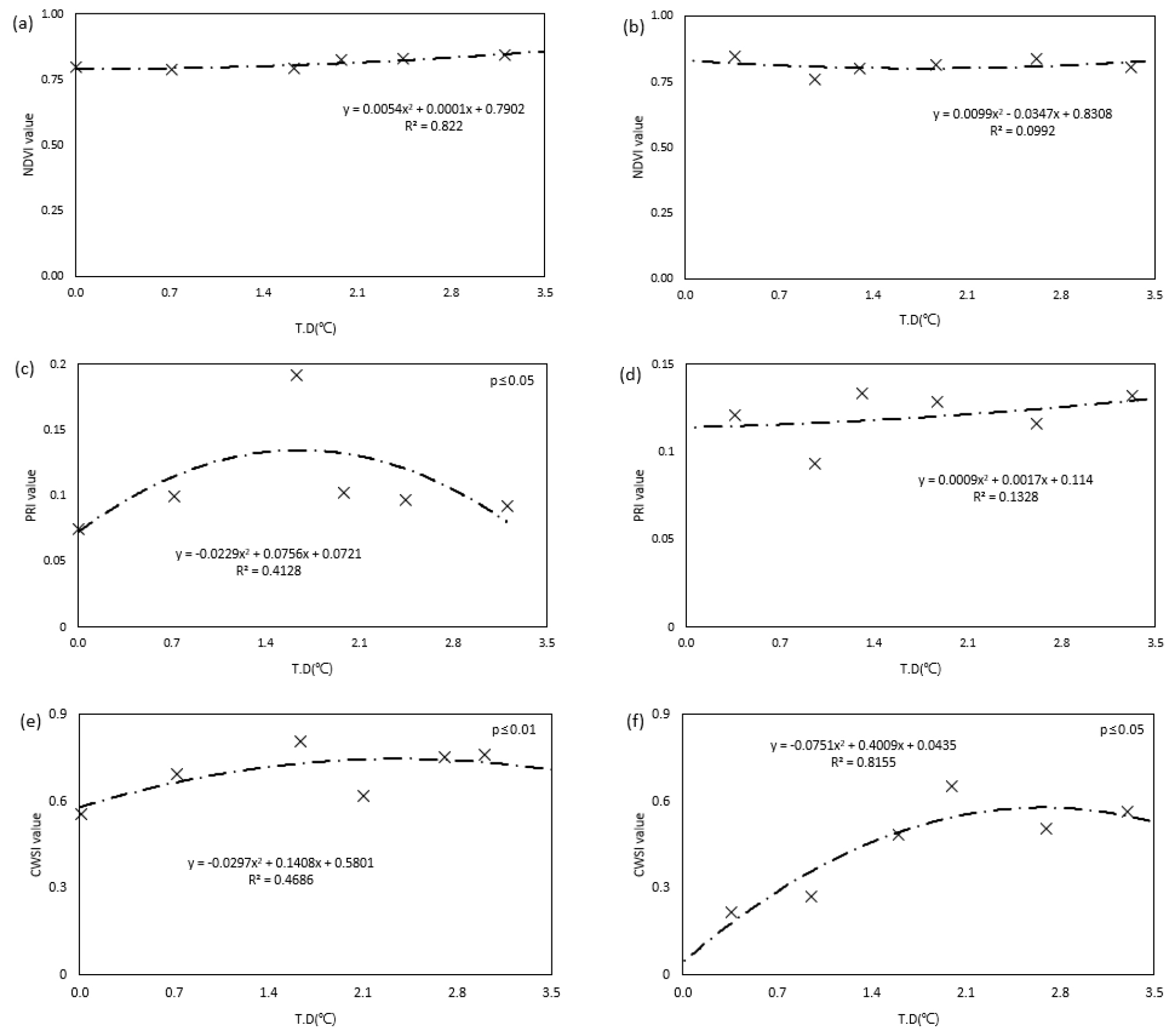
Fig. 7.
Comparison of the average values of each vegetation index ((a): NDVI in Sumi, (b): NDVI in Jopung, (c): PRI in Sumi, (d): PRI in Jopung, (e): CWSI in Sumi, (f): CWSI in Jopung) according to temperature treatment during the tuber enlargement period and peak growth period that determines potato yield. * T.D (°C): Temperature Difference compared to external conditions.
고온스트레스 시 생육변화 분석을 위한 최적 분석방법 탐색
고온에 따른 생육변화의 결과를 토대로 조풍보다 수미가 고온 스트레스를 많이 받는 품종이라는 점을 알 수 있었다. 이에 수미를 대상으로 고온에서 생육변화를 효율적으로 분석할 수 있는 방법을 알아보기 위해 생육 및 식생지수 값을 대조구(외부와 같은 온도) 및 고온 조건(3.2℃ 상승한 온도)에서 비교 분석하였다(Fig. 8). 고온 조건 시 초장은 증가하였으며 엽수는 생육 초기에는 더 빨리 증가하였으나 파종 후 50일경 이후부터는 고온 조건에서 감소하는 경향을 보였다. SPAD는 재배기간 중 감소하는 경향을 보였는데 고온 조건에서 감소되는 폭이 더 컸으며 NDVI는 고온 조건에서 생육이 왕성해지면서 더 높은 수치를 보였다. PRI는 고온 조건이 대조구에 비해 값의 범위가 높은 값이었지만, 고온으로 인한 엽록소 감소(Fig. 8c)로 생육 후기로 갈수록 감소하는 경향이었고, 대조구의 PRI 값은 비슷한 값을 유지하였다. PRI는 광합성 색소 중 제아크산틴의 변화를 나타내어 광합성 효율을 확인할 수 있는 지표로, 광합성 효율에 따른 최종 산물인 괴경 수량을 추정할 수 있는 도구가 될 수 있다(Romero et al., 2017). 특히 PRI는 생육후기의 엽록소 함량이 변화하는 시기 동안의 열 스트레스에 민감하다고 하는데, 이는 작물이 오랜 기간동안 고온스트레스를 받을 때 나타나는 엽록소 함량 감소, NPQ활성 감소 등에 따른 광합성 효율 감소에 따라 PRI값이 감소하기 때문이다(Cao et al., 2019). 따라서 PRI 값이 감소하게 되면 탄소대사 또한 감소되어 결국 괴경으로 축적되는 전분의 양이 감소되기 때문에 생산량 감소가 되는 것으로 진단할 수 있다(Hancock et al., 2014). CWSI는 파종 후 60일부터 고온 조건에서 높은 수치를 보이기 시작하여 괴경이 비대되는 시기인 63일 이후부터 고온이 대조구 온도보다 상대적으로 높은 값을 보이는 것을 확인하였다. 이를 통해 고온에서 생육조사가 필요 없이 쉽게 감자의 생육을 진단하기 위해서는 3개의 센싱기술 중 PRI센서를 이용한 분석이 가장 용이하게 사용될 수 있을 것으로 판단된다.
이는 고온 스트레스로 인해 작물의 생산량과 품질의 저하가 예상되는 기후변화 시나리오에서 생육단계 중에서도 수확량을 결정지을 수 있는 괴경 비대기에 고온이 다양한 생육요소에 미치는 영향을 조기진단하고, 이를 통해 수확량을 예측할 수 있는 잠재력을 제시한다.
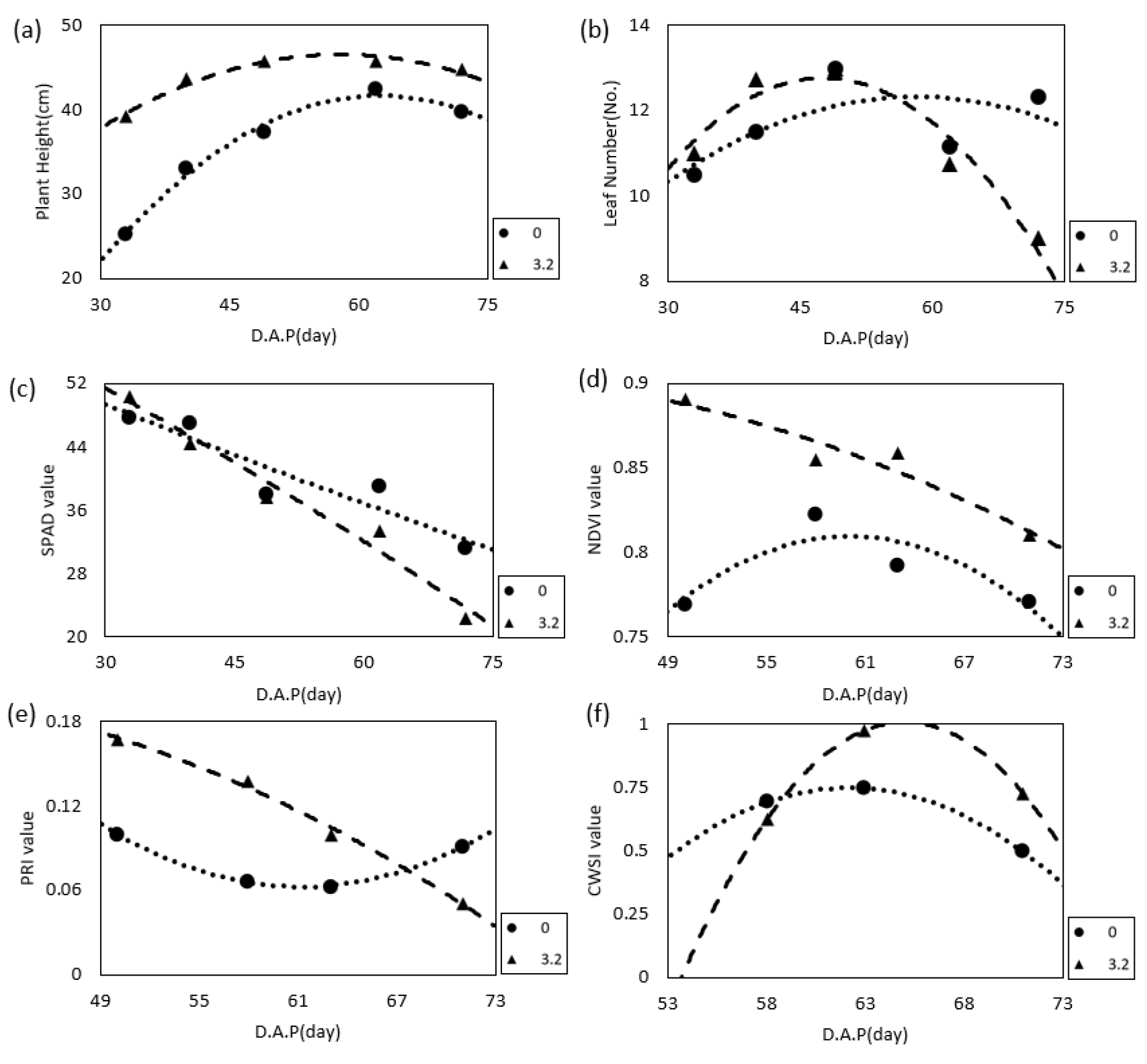
Fig. 8.
Comparison of growth responses((a): Plant Height, (b): Leaf Number, (c): SPAD, (d): NDVI, (e): PRI, (f): CWSI) in high-sensitive Sumi variety between control (0°C) and high temperature treatment (+3°C), which is more susceptible to high temperature between the two varieties. * D.A.P (day): Day After Planting.
적 요
기후 온난화에 대응하여 온도구배하우스에서 온도가 상승한 정도를 하우스 입구로부터 거리별로 6구간으로 나누어서 관찰하였을 때 두 품종에서 나타나는 생육, 수량 및 식생지수 변화를 분석한 결과는 다음과 같다.
1.수미는 1.6℃ 상승한 온도에서 지상부의 생육과 SPAD값이 감소되며 2.4℃ 이상의 온도가 상승했을 때 수량이 감소하였다.
2.조풍에서는 3.5℃ 이하로 온도가 상승하는 경우 고온, 생육, 수량 간의 경향성이 나타나지 않아 수미에 비해 고온에 적합한 품종으로 판단된다.
3.수미를 대상으로, 대조구(하우스 외부와 동일한 온도)와 고온 조건(3.2℃ 상승한 온도)의 생육 반응들을 비교해보았을 때, 초장은 고온 조건에서 높은 값을 유지하였고, 엽수는 고온조건에서는 빠르게 증가하다가 파종 후 49일째부터 감소하는 경향을 보였으며 대조구에서는 점차적으로 증가하는 경향을 보였다. SPAD는 파종 후 30일에 모두 감소하는 경향을 보였는데, 고온조건에서 더 빠르게 감소하는 경향을 보였다.
4.지구온난화로 인한 온도 상승 시 수미의 생육 적정 온도에 대한 한계점은 외부온도 대비 1.6℃ 상승한 경우였으며, 센서 중 PRI가 생육 및 수량과 유사한 경향을 보인 것을 확인할 수 있었다.
