

Maize is one of the world’s three major crops with rice and wheat. In Korea, maize cultivation is front to abiotic stresses such as drought at early growth stage, wet in middle growth stage, and chilling stress at heading stage. The drought and hot air stresses are most damaging to biomass production, which most actively progresses before a rainy period so called monsoon. Therefore, a breeding endeavor for the selection of hot air tolerant maize lines was further needed. In the climate condition of Korea, maize is cultivated during dry season at least oveR 2 months during the young vegetative stage. Therefore, the effective selection of drought-tolerant maize lines is highly important in the Republic of Korea. Biomass production and survival consequence are closely related to photosynthetic activity under dry and hot conditions.
In the context of hot air stress, Chlorophyll a fluorescence transient analysis (so-called JIP-test), and chlorophyll fluorescence imaging (CFI) technique may be applied to investigate the energetic behaviour of photosynthetic sensory systems. The JIP-test is a tool to analyse the polyphasic rise of the Chlorophyll a (Chl a) fluorescence transients (phases labelled “OJIP”). Although it corresponds to only a very small fraction of the dissipated energy from the photosynthetic apparatus of fruit surface, Chl a fluorescence is widely accepted to provide a means to a better understanding of the structure and function of the photosynthetic apparatus. At room temperature, the Chl a fluorescence of plants, algae, and cyanobacteria in the 680-740 nm spectral region is emitted mainly by photosystem (PS) II, and thus it can serve as an intrinsic probe of the fate of its excitation energy. The spectra and the kinetics of Chl a fluorescence are powerful and non-invasive tools for such investigations. The primary use of fluorescence has included the estimation of chlorophyll concentration and pigment-protein interactions, and studies on the stability of thylakoid membranes. However, the relationship between chlorophyll and in vivo fluorescence varies widely over time and space. These processes include species changes, nutrient concentrations, and incident radiation (Falkowski and Raven, 2013).
Most studies analysing the effects of heat or chilling stress on OJIP transients have been conducted on plant leaves (Ogweno et al., 2009; Wahid et al., 2007) but not precisely in maize leaves. Even these studies have been limited to apple (Chen and Cheng, 2009; Chen et al., 2009, 2008). Photosynthetic activities differ between leaves and fruits; for example, in the pericarp of cherry tomato, photosynthetic fixation of 14CO2 has been shown to occur at higher rates than in the leaves(Laval-Martin et al., 1977).
The photosynthetic apparatus is the most sensitive component in evaluating the degree of temperature-related stress damage (Rachmilevitch et al., 2006). CFI technique has been mainly used as an effective tool in order to study the damage and activity of the electron transport in the photosynthetic system under various environmental stresses. CFI as a rapid and non-destructive technique has quickly progressed, and has been used successfully in evaluating plant photosynthetic activity (Yoo et al., 2014). This study was performed to evaluate the validity of CFI technology to determine the degree of hot air stress in maize leaves.
MATERIALS AND METHODS
Growth Condition of Plants
Maize (Zea mays L.) varieties were Kwangpyeongok and Ilmichal. Seeds were sown in pots mixed with basal N-P-K (12-5-8 kg/10a). When the third leaves sprang, the pots were transferred to a growth chamber (50 days). The rapid hot air was performed for 96 hours (12 hours 2,500 μmol m-2 s-1 illumination of the 24 hours). All of the treatments were prior to exposure to hot air condition (35±2°C). Maize seedlings were placed in a chamber and kept at room temperature. The growth chamber was set to 35% relative humidity. All treatments were performed and each treatment was carried out in triplicate.
Measurement of Chlorophyll Fluorescence Imaging The maize seedlings were measured partially for each treatment after treatment of the hot air stress. A CFI fluorcam (Handy FluorCam FC 1000-H, PSI, Czech Republic) was used to measure the fluorescence images of the leaves.
The source of actinic light was orange LED at an intensity of 200 m-2 s-1. The source of saturating light was a halogen lamp with an intensity of 2,500 m-2 s-1. The fluorescence parameters maximum quantum efficiency of PS II (Fv/Fm), PS II operating efficiency (ΦPS II = F'q/F'm), and non-photochemical quenching (NPQ) were monitored by quenching kinetics analysis (Barbagallo et al., 2003; Genty et al., 1989; Genty et al., 1990). The data were calculated according to the parameters of the CFI fluorCam, which measured quenching kinetic. The light conditions were actinic light, red LED, 200 μmol m-2 s-1, and saturating light, moderate light, 1,250 μmol m-2 s-1.cence Parameters
Chlorophyll fluorescence parameters were defined as follows (Gorbe and Calatayud, 2012):
F0: Minimal chlorophyll fluorescence intensity measured in the dark-adapted state when all PS II RCs are open
Fm: Maximal chlorophyll fluorescence intensity measured in the dark-adapted state during the application of a saturating pulse of light
Fp: Maximum fluorescence value under saturating illumination when all reaction centers are closed or in reduced state
Fv: Variable chlorophyll fluorescence (Fm− F0) measured in the dark-adapted state when non-photochemical processes are minimum
Ft: Fluorescence at time t after onset of actinic illumination
Fv/Fm: Maximum quantum yield of PSII photochemistry measured in the dark-adapted state
ΦPSII: Effective quantum yield of photochemical energy conversion in PS II (F'q/F'm)
NPQ: Nonphotochemical quenching
Rfd: Fluorescence decrease ratio. In this case, Fs is obtained after illumination with continuous saturating irradiance. Rfd is an indicator of the photosynthetic quantum conversion and is correlated with CO2 fixation rates
RESULTS AND DISCUSSION
Chlorophyll Fluorescence Imaging (CFI) Analysis
Maize seedlings were apparently similar. Thus, it was not easy to determine the damage on the eyes before 36 hours after hot air (Fig. 1). In hot air stressed Kwangpyeongok and Ilmichal leaves, dark-adapted fluorescence value (red color) at Fo was lower than in control at 24 hours after treatment. In case of Kwangpyeongok, Fm value of 2,962 was about 8.5 fold lower in control than in hot air stressed leaves of 344. Ilmichal Fm value of 2,753 was about 2.8 fold lower in control than in hot air stressed leaves of 993 (Fig. 2). CFI was initiated to investigate whether it can be used as a reliable indicator to evaluate the hot air stress of maize seedlings in order to study the light stress-specific difference that may be involved in the photosynthetic apparatus. CFI has been applied for different purposes in the postharvest life of fruits (Chen et al., 2009), but the main focus has been on detecting factors that can increase or decrease product quality (Yoo et al., 2014). Moreover, the technique has been used for various objectives both in pre- and post-harvest conditions for the detection of biotic or abiotic stresses in plants and plant products. In comparison to the application of CFI for the detection of abiotic stresses in two maize cultivars (Chen et al., 2009; Gorbe and Calatayud, 2012), this study focused on the photochemical responses to hot air stress condition. At least, CFI technique is applicable to determine the hot air stress in numeric transformation of direct measured fluorescence values (Fig. 2)

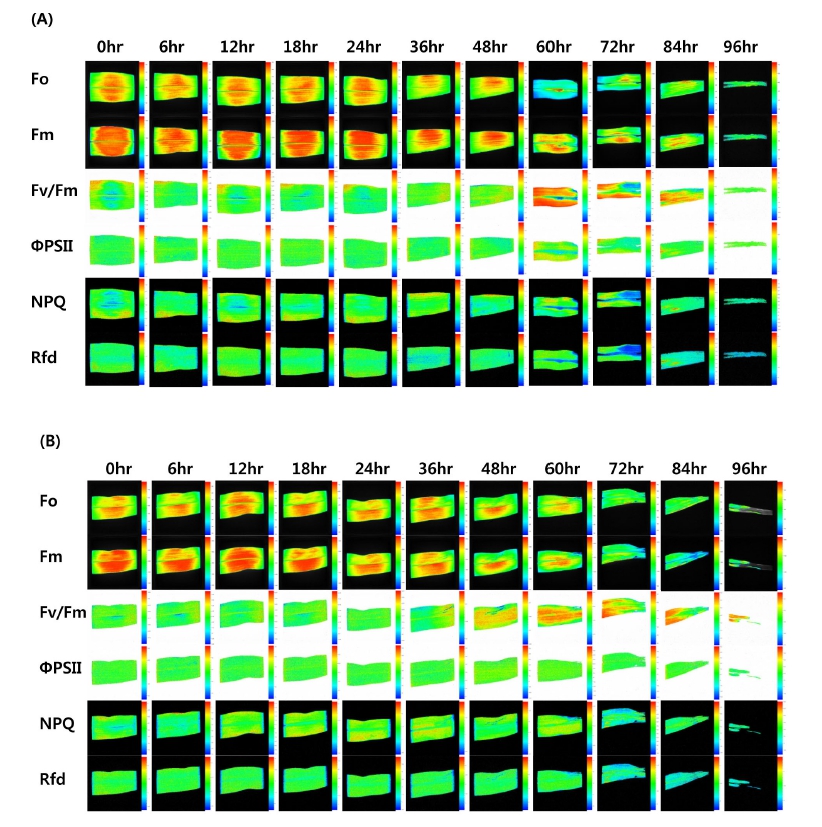
Maximum Quantum Yield (Fv/Fm) and Other Parameters
The maximum quantum yield (Fv/Fm) was 0.77 (Kwangpyeongok) and 0.78 (Ilmichal) before hot air stress. It was not changed until 36 hours. Although the Fv/Fm ratios in hot air stress maize leaves were almost normal, Fm and Fv values were far lower than in control seedling (Fig. 3A, B). Also, maximum fluorescence value (Fp) under saturating illumination when all reaction centers are closed or in reduced state was extremely lowered by hot air stress. Ft values were considerably reduced already at 12 hours. In hot air stressed leaves, all measured fluorescence parameters declined to over half compared to control seedling.
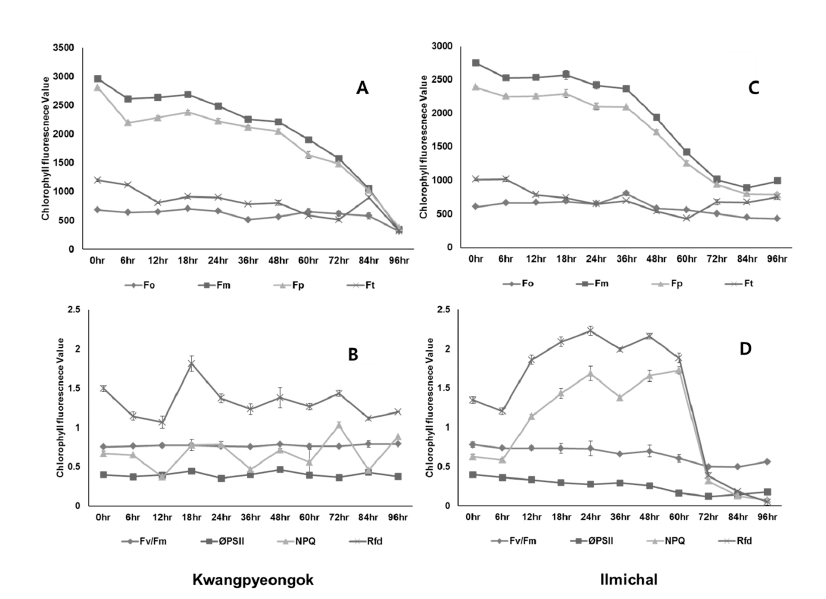
Fig. 3.
Comparison of changes in chlorophyll a fluorescence imaging parameters from maize seedlings grown in growth chamber maintained at 35±2°C with 35% relative humidity for 96 hr under hot air condition. Comparative analysis of Fo, Fm, Fp and Ft (3A and C) and Fv/Fm, ΦPSII, NPQ, and Rfd (3B and D). *A and B : Kwangpyeongok , C and D : IlmichalEffective Quantum Yield of Photochemical Energy Conversion (ΦPS II)
In hot air stressed maize leaves, the ΦPS II has decreased from 0.38 to 0.04 at 96 hours, indicating that the effective quantum yield of photochemical conversion in PSII was inactive (Fig. 3B and D). At 48 hours, Fv/Fm value was not changed from 0.77 to 0.74 under hot air stress in Kwangpyeongok and at 24 hours, Fv/Fm value was not changed from 0.78 to 0.73 under hot air stress in Ilmichal. It may mean that the maize leaves are still photosynthetically active. Therefore, this photochemical parameter cannot be a good indicator for the determination of hot air stress. In general, healthy plants have a very conservative Fv/Fm value of about 0.8 (Baker, 2008). In hot air stress condition, Fv/Fm was greatly retained compared to control. Björkman and Demmig (1987) and Johnson et al.(1993) also reported optimal values of Fv/Fm around 0.8 for most plant species, and values lower than this are observed in plants exposed to stress, indicating in particular the phenomenon of photoinhibition. However, it is still unclear whether hot air stress is the case.
In hot air stressed maize leaves, Ilmichal ΦPSII gradually declined from 0.39 to 0.12 within 72 hours and Kwangpyeongok ΦPSII also gradually declined from 0.38 to 0.26 within 72 hours (Fig. 3). This implies that they were photosynthetically inactive. The lower ΦPSII values in hot air stressed maize leaves were assumed to be low Chlorophyll A florescence in steady state; i.e., under continuous light pulse. In general, the chlorophyll fluorescence neaR 25°C is most sensitive as the temperature in the growth chamber used in this study (Baker, 2008; Baker and Rosenqvist, 2004; Fracheboud et al., 1999). However, it was thought that this lower effective quantum yield of photochemical energy conversion in PS II was related to lower electron transport activity to Q-A resulting in severe stress on electron-transport from PSII to PSI.
Non-photochemical Quenching (NPQ)
In hot air stressed leaves, the Kwangpyeongok NPQ value has steeply increased from 0.51 to 1.28 at 72 hours (Fig. 3B). However, the final value was enhanced down to 0.02 at 96 hours, and the Ilmichal NPQ value has steeply increased from 0.62 to 1.72 at 60 hours (Fig. 3D). However, the final value was enhanced down to 0.08 at 96 hours.
The NPQ values in hot air stressed Kwangpyeongok leaves have gradually increased from 1.34 to 1.90 at 72 hours and Ilmichal leaves also increased from 0.62 to 1.72 at 60hours. In general, the increase in NPQ means inefficiency and energy dissipation in photosynthetic apparatus (Genty et al., 1990; Havaux et al., 1991; Johnson et al., 1993). It may imply that the NPQ parameter is most sensitive to hot air stress because the higher value of NPQ indicates a low possibility of photosynthetic electron transport resulting in an increase in the inactive chlorophyll of maize leaves (Genty et al., 1989; Havaux et al., 1991). Thus, these NPQ values of maize leaves can be an available indicator parameter
Fluorescence Decrease Ratio (Rfd)
The Rfd value was considerably increased from 1.34 to 1.83 at 12 hours, and then decreased to 0.17 at 96 hours in Kwangpyeongok. At 72 hours, the Rfd value was again increased to 1.90. In Ilmichal the Rfd value also increased from 0.62 to 1.72 at 60 hours and then decreased to 0.08 at 96 hours (Fig. 3D).
In an earlier report (Krupa et al., 1993), the decrease of fluorescence decrease ratio (Rfd) in tomatoes under the sprinkler irrigation could be explained as photoinhibitory effect on the Calvin cycle by changes in the environmental conditions such as temperature, oxygen, light, and physiological status of plants. There is no report until now on the Rfd value under hot air stress. Although the increase in Rfd seems to be hot air stress as shown in Fig. 3B, the stressed maize leaves did not accompany the increase of ΦPSII. Thus, it could be assumed that the stressed leaves did not recover to normal status. When this parameter uses as a stress indicator, therefore, it should be carefully applied
The hot air condition seemed to cause severe damage to the photosynthetic apparatus, resulting in changes in the photosynthetic activity of maize leaves. The measurement of photochemical responses may be able to determine the photosynthetic stress of maize leaves under hot air.
This study has shown that CFI can be used as a reliable tool to evaluate the healthy condition of maize leaves.
In maize leaves exposed to continuous hot air stress condition for night, NPQ values increased gradually until 60 hours. NPQ values in steady state have nonphotoquenching characteristics in dark-adapted state (Baker, 2008). Although changes in NPQ are nonlinearly related to higher values than ΦPSII in leaves as earlier suggested (Baker, 2008, 1991), the ΦPSII seemed to be applicable to determine the stress under hot air condition.
The hot air stress causes severe damage to the photosynthetic apparatus, resulting in changes in the apparent viability of leaves. This study has clearly shown that CFI can be used as a reliable tool to evaluate the hot air stress of maize leaves.
CONCLUSIONS
The fluorescence imaging and the numeric data of Fv/Fm, ΦPSII, and NPQ showed that different responses occurred under hot air stress condition. This practical study of the CFI technique has shown that the numeric values in Fv/Fm, ΦPSII, and NPQ could be applied to determine the hot air stress. Specifically, Fm and ΦPSII values declined under hot air stress. Fv/Fm values retained almost 0.8. On the basis of these results, the photochemical numeric parameters obtained from CFI were useful for an evaluation of hot air stressed maize leaves. The numeric data derived from CFI analysis can be a rapid method for hot air stress determination.
