

서 론
재료 및 방법
재배방법 및 처리내용
생육조사 및 데이터 측정
주요 변수 선택
랜덤 포레스트 분류 모델
통계분석
결과 및 고찰
벼 생육 분석
엽록소 농도 및 SPAD 분석
엽분광 데이터 분석
주요 파장 선택 및 모델 성능 평가
적 요
서 론
질소는 엽록소를 구성하는 성분으로 광합성과 직접적으로 관련이 있으며 작물의 생육과 생산량에 중요한 영향을 미치는 필수 요소 중 하나이다(Peng et al., 2021). 질소 시비가 작물 생육 증진과 생산량 증가에 미치는 효과는 여러 연구에서 보고되었다(Amanullah et al., 2016; Hammad et al., 2022).
하지만 90년대 이후 질소 비료 사용량이 증가하였고 질산염 오염, 아산화질소 등 질소 산화물과 암모니아는 환경 오염을 유발하고 있다(Byrnes, 1990). 특히 온실가스의 하나인 아산화질소(N20)는 전체 배출량의 약 84%가 농업 분야에서 발생하며, 그 대부분은 질소질 비료 사용에 의해 발생한다(Song et al., 2021). 이러한 상황에서 탄소중립, 친환경 농법 등 지속 가능한 농업의 중요성이 부각되면서, 적정량의 비료 시비를 위해 재배지의 양분 상태, 시비 수준을 평가하는 기술의 필요성이 높아지고 있다.
작물의 영양상태는 전통적으로 켈달법(Kjeldahl method)과 듀마스법(Dumas method) 같은 유기체 질소 함량 분석법을 통해 파괴적으로 측정하였다. 켈달법과 듀마스법은 질소 함량을 정확하게 측정할 수 있지만, 분석 시간이 오래 걸려 실시간 관측이나 노지 현장 적용이 어렵다는 한계가 있다. 최근 센싱 기술의 발전으로 인해 비파괴적인 방법으로 작물의 생육과 영양 상태를 측정하려는 다양한 연구들이 활발히 수행되고 있다. 특히 다분광, 초분광 센서를 통해 측정되는 분광 정보는 식물의 구조적, 생리적 상태에 따라 특정 파장 영역의 흡광도, 반사도, 투과율이 달라지기 때문에 작물의 생육 상태를 파악하는데 활용되고 있다(Sishodia et al., 2020). 접촉식 분광 센서인 엽록소 측정기(SPAD-502, Konica Minolta)는 적색과 적외선 영역의 흡광도 비율을 이용하여 SPAD (Soil Plant Analysis Development) 값을 측정한다. SPAD 값은 잎의 엽록소 농도와 높은 상관성을 보여, 비파괴적으로 엽록소 농도를 측정하고 작물의 질소 상태를 평가하는데 이용되고 있다(Markwell et al., 1995; Uddling et al., 2007; Wu et al., 2024; Yuan et al., 2016). 또한 특정 파장대역의 분광정보를 활용하여 계산되는 식생지수(Vegetation Index)는 작물의 생육 상태를 정량화하기 위해 개발된 지수로, 드론이나 인공위성 영상 등 원격 센싱을 통해 작물의 생육을 분석하는데 사용되고 있다(Bannari et al., 1995). 하지만 SPAD 값은 적색과 적외선 영역의 제한된 파장을 사용하기 때문에 엽록소 외에 카로티노이드, 안토시아닌 등 다른 색소가 흡수하는 광 영역의 영향을 분석하는데 한계가 있다. 식생지수 역시 제한된 파장 영역을 이용하여 계산되며 이를 보완하기 위해 사용 목적에 맞춘 다양한 지수가 개발되었다. 그러나 드론이나 위성 등 비접촉식 센서와 영상은 측정 지역의 토양 반사율, 광조건, 대기 상태의 영향을 받아 정확도가 떨어질 수 있다(Gai et al., 2024).
접촉식 엽분광계는 식물의 잎에 센서를 접촉시켜 흡광도, 반사율, 투과율을 측정하는 장비로 가시광선부터 근적외선 영역까지 비파괴적으로 식물의 광학적 특성을 조사할 수 있다. 외부 환경의 영향을 적게 받으며, 1 nm 단위로 측정이 가능해 특정 밴드 값을 활용하여 SPAD 값이나 식생지수를 계산할 수 있다. Oliveira et al. (2017)은 엽분광계를 사용하여 유칼립투스 잎의 반사율을 측정하고 식생지수를 계산하여 잎의 질소 농도를 추정하였으며 특정 식생지수(reNDVI, mNDI)가 질소 농도와 높은 상관관계를 나타내는 것을 확인하였다(r=0.97). 또한 Gai et al. (2024)은 엽분광계를 이용하여 한발스트레스에 의한 사탕수수의 엽록소 함량 변화를 분석하였으며 반사율 데이터를 통해 엽록소 함량을 추정하였다(R2=0.98).
따라서 본 연구에서는 재배현장에서 간단하고 비파괴적인 방법으로 포장의 시비 수준을 평가하여 작물의 상태를 진단하기 위해 엽분광계를 이용하여 질소 시비 수준이 다른 포장에서 재배한 벼의 생육시기별 분광 반사율을 측정하고 이를 기반으로 질소 시비 수준 분류를 위한 머신러닝 모델을 학습시켜 성능을 평가하였다.
재료 및 방법
재배방법 및 처리내용
본 연구는 2022년 전라북도 완주군에 위치한 국립식량과학원 내 최적화 온실에서 수행하였으며 품종은 신동진 벼를 사용하였다. 5월 중하순에 파종하여 6월 초순에 30 × 14 cm 간격으로 주당 3~4본을 이앙하였다.
질소 시비는 요소비료를 사용하였으며 표준 질소 시비량(9 kg/10a)을 기준으로 무비(0%), 소비(50%), 표준시비(100%), 과비(200%) 4수준으로 처리하였고 기비, 분얼비, 수비를 5:3:2의 비율로 나누어 시비하였다. 칼륨은 염화가리를 기비, 수비 5:2의 비율로 분시하였고 인산은 용성인비를 전량 기비로 시비하였으며 표준 시비량에 따라 전체 처리구에 동일한 양을 시비하였다. 시험 전 전체 토양을 경운하고 슬레이트 판을 설치하여 8개의 처리 구획을 만들었으며, 인접한 처리구와 일사 방향에 의한 영향을 최소화하기 위해 서편에 위치한 처리구획은 북에서 남쪽 방향으로 질소 시비 수준이 증가하도록 처리하였고, 동편에 위치한 처리구획은 반복구로 설정하고 역방향으로 배치하였다.
생육조사 및 데이터 측정
질소 시비 수준에 따른 생육 차이를 확인하고자 처리구마다 5개체를 지정하여 7월 중순부터 9월 중순까지 2주 간격으로 초장과 분얼수를 조사하였으며 SPAD-502 를 이용하여 신엽 바로 아래 잎을 대상으로 SPAD 값을 측정하고 실제 엽록소 함량과 비교를 위해 동일 처리구에서 조사 대상이 아닌 개체의 잎을 채취하여 엽록소 a, 엽록소 b를 포함한 엽록소 함량과 카로티노이드 함량을 분석하였다.
이앙 후 30일경부터 엽분광계(CI-710s, Bio-Science)를 사용하여 잎의 분광 반사율(%)을 측정하였다. 기기는 매뉴얼에 따라 흑백 반사판을 이용하여 보정(calibration)한 후에 사용하였으며, 7~10일 간격으로 맑은 날 오전 9시에서 12시 사이에 각 처리구마다 10개체를 지정하여 신엽 바로 아래 잎을 대상으로 3회 반복 측정하였다. 엽분광계로 측정한 파장 영역 중 380~949 nm 영역을 분석에 사용하였으며 삼차 보간법(cubic interpolation)을 통해 1 nm 간격으로 반사율을 보정하여 3회 반복 측정한 평균값을 사용하였다.
주요 변수 선택
분광 반사율 데이터는 인접한 파장 간 상관관계가 높기 때문에 다중 공선성 문제가 발생하며 분류나 예측 모델이 과대적합(overfitting) 되는 원인이 될 수 있다(Wei et al., 2021). 이를 해결하기 위해 특정 밴드를 선택하여 식생지수를 계산하거나 주성분회귀(Principal Component Regression; PCR), 부분최소제곱법(Partial Least Squares; PLS) 등을 이용하여 중요 변수를 추출하는 방법 등이 사용되었다(Hansen & Schjoerring, 2003; Li et al., 2020). 본 연구에서는 모델의 입력 변수를 선택하기 위해 측정시기별로 분광 반사율과 반사율 미분 값의 처리 내 분산과 처리 간 분산을 비교하여, 처리 간 분산이 더 큰 파장을 추출하였다. 또한 추출된 파장들 간의 상관관계를 분석하여 상관성이 높은 파장들을 그룹별로 묶고, 부분최소제곱법 분석을 통해 모델 변수로 사용될 주요 파장을 선택하였다. 부분최소제곱법 분석은 파이썬(Python)의 사이킷 런(scikit-learn) 라이브러리를 사용하였으며 선형 결합 개수(n_component)는 그룹 내 파장 개수의 30%로 설정하였다. 모델은 100회 반복하여 실행하였고 각 파장에 대한 평균 Variable Importance in Projection(VIP) 값을 계산하였다. 이후 각 측정시기별로 VIP 값이 높은 파장을 선택하여 25개의 변수를 모델에 사용하였다.
랜덤 포레스트 분류 모델
본 연구에서는 무비, 소비, 표준시비, 과비의 4가지 질소 시비 수준을 분류하기 위해 사이킷 런의 랜덤포레스트 분류 모델(RandomForestClassifier)을 활용하였다. 측정시기별로 선택된 주요 파장의 반사율을 변수로 사용하였으며, 전체 측정데이터를 표준화(StandardScaler)한 후, 학습 데이터와 평가 데이터를 7:3의 비율로 분할하였다. 트리(tree) 개수는 100개로 설정하였으며, 각 측정시기별 트리의 최대 깊이(max depth)는 21로 설정하여 모델을 학습을 학습하였고, 혼돈행렬(Confusion matrix) 값을 바탕으로 학습된 모델의 각 측정시기별 분류 정확도(Accuracy)를 계산하고 성능을 비교, 평가하였다(Table 1).
Table 1.
Confusion matrix and performance metrics.
* TP, TN, FP, FN denote true positive, true negative, false positive, and false negative, respectively.
* Precision is defined as the ratio of correctly predicted positive observations to the total predicted positives.
통계분석
초장, 분얼수, 엽록소농도, SPAD 값의 처리 간 비교를 위해 유의수준 5%에서 일원분산분석(One-way ANOVA)을 실시하였으며, 유의한 차이가 있는 경우 최소 유의차 검정(LSD)을 수행하였다.
결과 및 고찰
벼 생육 분석
질소 시비 수준에 따른 벼의 초장과 분얼수를 비교한 결과 생육 초기에 측정된 분얼수를 제외하고 전 생육기간 동안 질소 시비량이 증가할수록 초장과 분얼수는 증가하였다(Fig. 1). 수확 전 초장을 조사했을 때 무비에 비해 소비는 11%, 표준시비는 21%, 과비는 32% 증가하였으며 분얼수는 소비는 2개, 표준시비는 3개, 과비는 4.8개 많았다(Table 2). 질소 시비 수준에 따른 초장과 분얼수의 변화로 질소시비처리로 생육 증진 효과가 있음을 확인하였다.
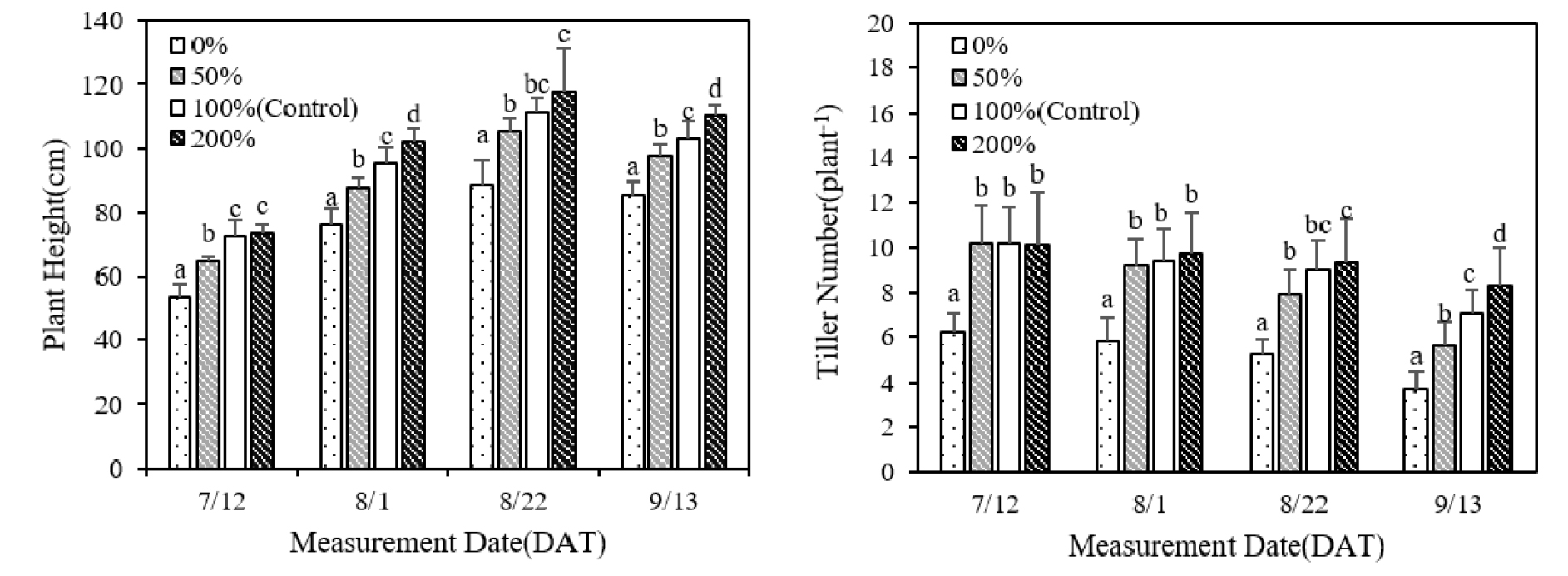
Fig. 1.
Comparison of plant height and number of tillers at different stages of growth in rice plants subjected to different levels of nitrogen fertilization (p < 0.05). (* For measurements performed on the same date, the same lower-case letters above bars indicate no significant difference between treatments at the p < 0.05 level, as determined using the least significant difference test.)
Table 2.
Comparison of plant height and number of tillers at different stages of growth in rice plants subjected to different levels of nitrogen fertilization.
엽록소 농도 및 SPAD 분석
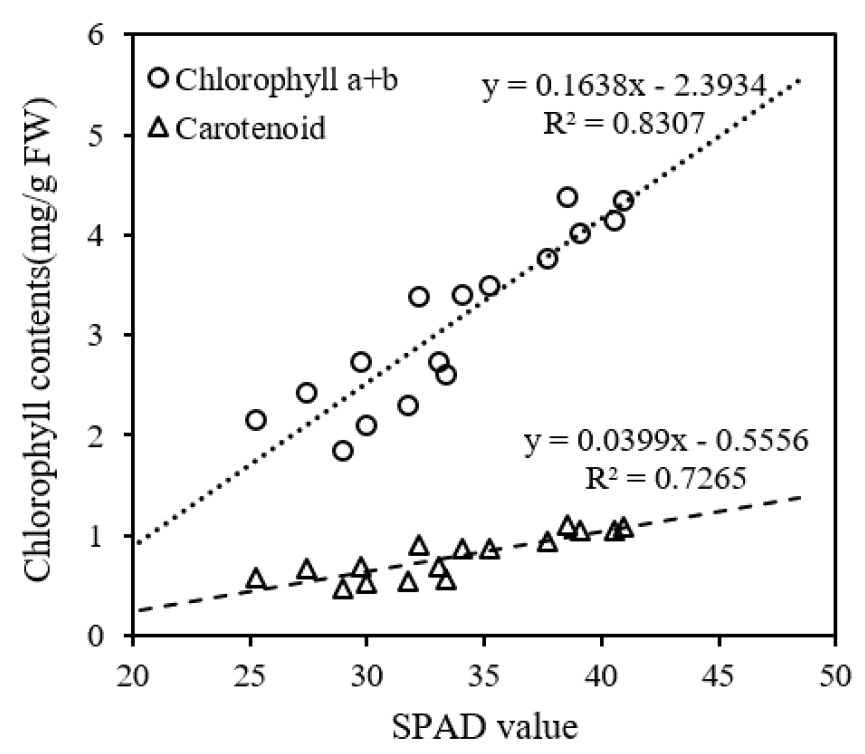
질소시비량이 증가할수록 잎의 엽록소와 카로티노이드 농도가 증가하였으며 측정시기에 따른 변화를 분석한 결과 모든 처리에서 8월 초순까지 농도가 감소하였고 두번째 측정 이후 증가하였다가 8월 하순부터 다시 감소하였다(Fig. 2). 또한 질소시비량이 증가하면 SPAD 값이 증가하였으며 7월 중순과 8월 초순에 무비, 소비 처리의 SPAD 값은 표준시비, 과비 처리와 차이가 있었고 8월 하순에는 각 처리 간 유의한 차이가 있었다(Fig. 3). 측정시기별 변화 양상은 엽록소 농도와 유사한 경향을 보였으며 엽록소 농도와 SPAD 값간 양의 상관관계를 확인하였다(Fig. 4). Ghosh et al. (2020)은 분얼기에 잎의 질소 함량과 SPAD 값이 가장 높았으며, 출수 이후 생육이 진행되면서 이삭과 종실의 질소 요구량이 증가하여 잎의 질소 함량과 SPAD 값이 감소함을 확인하였으며 질소 시비 시점이 측정값에 영향을 주었다. 질소는 아미노산, 단백질, 핵산 등 식물 세포 구성 성분의 중요한 요소로 질소시비량이 증가하면 단백질 합성과 효소 활성이 촉진되며 엽록소와 카로티노이드와 같은 광합성 색소의 생성이 증가한다(Fathi, 2022; Taiz et al., 2015). 또한 유수 형성과 출수 이후, 이삭과 종실 형성에 필요한 질소는 잎과 다른 기관에 저장된 질소의 재분배를 통해 공급된다(Mae & Ohira, 1981; Tabuchi et al., 2007). 따라서 8월 초순에 분얼비로 시비된 질소는 소모되었고, 토양에서 공급되는 질소의 양이 감소함에 따라 유수 형성에 필요한 질소가 재분배되어 잎의 엽록소 농도와 SPAD 값이 감소한 것으로 보인다. 이후 수비를 시비한 후에는 토양으로부터 질소가 다시 공급되어 엽록소 농도와 SPAD 값이 증가한 것으로 판단되며, 8월 하순 이후에는 질소 재분배와 잎의 노화로 인해 엽록소 농도와 SPAD 값이 다시 감소한 것으로 보인다.
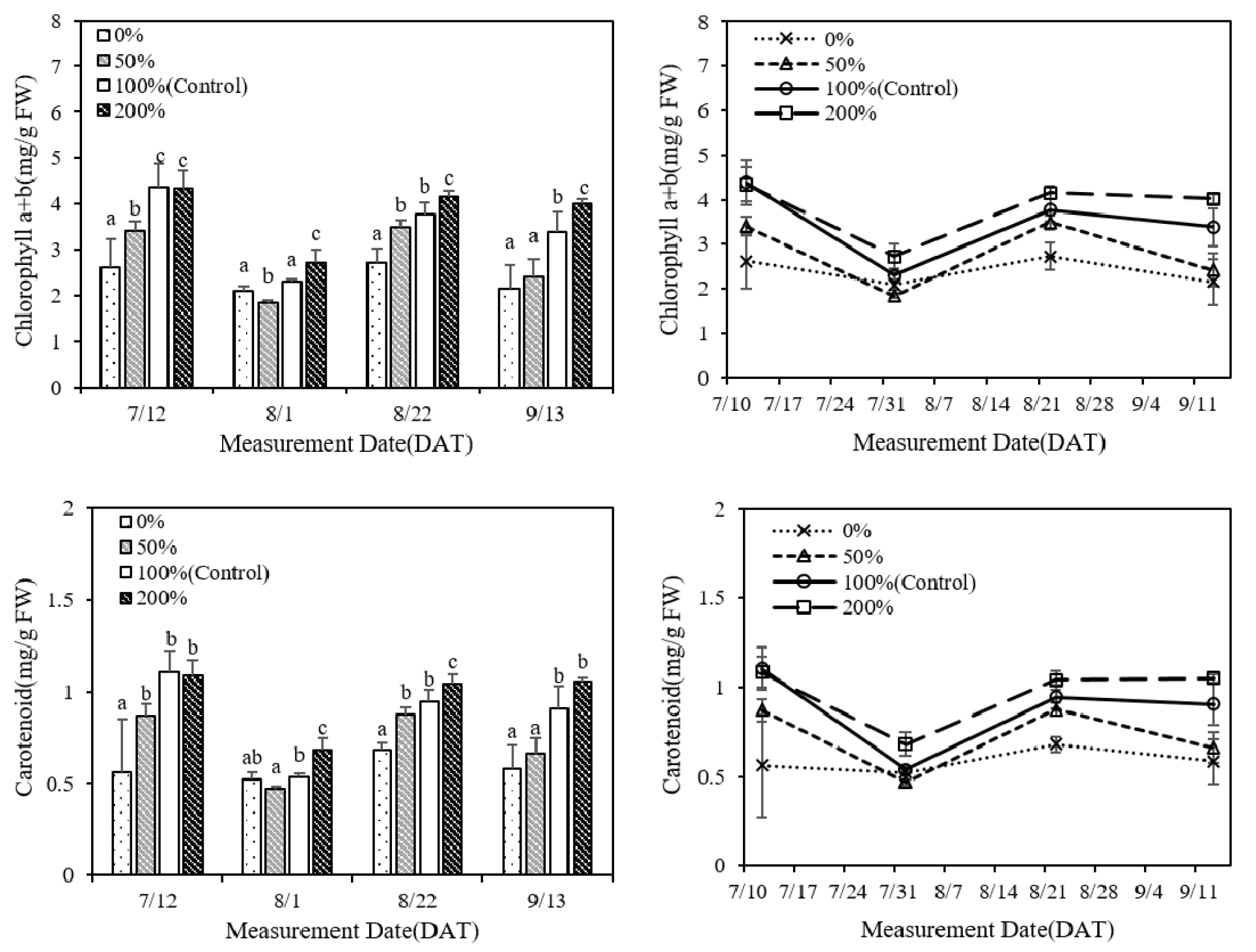
Fig. 2.
Comparison of leaf chlorophyll and carotenoid contents at different stages of growth in rice plants subjected to different levels of nitrogen fertilization (p < 0.05). (A) Chlorophyll (a + b) content, (B) Changes in chlorophyll (a + b) content, (C) Carotenoid content (D) Changes in carotenoid content. (* For measurements performed on the same date, the same lower-case letters above bars indicate no significant difference between treatments at the p < 0.05 level, as determined using the least significant difference test.)
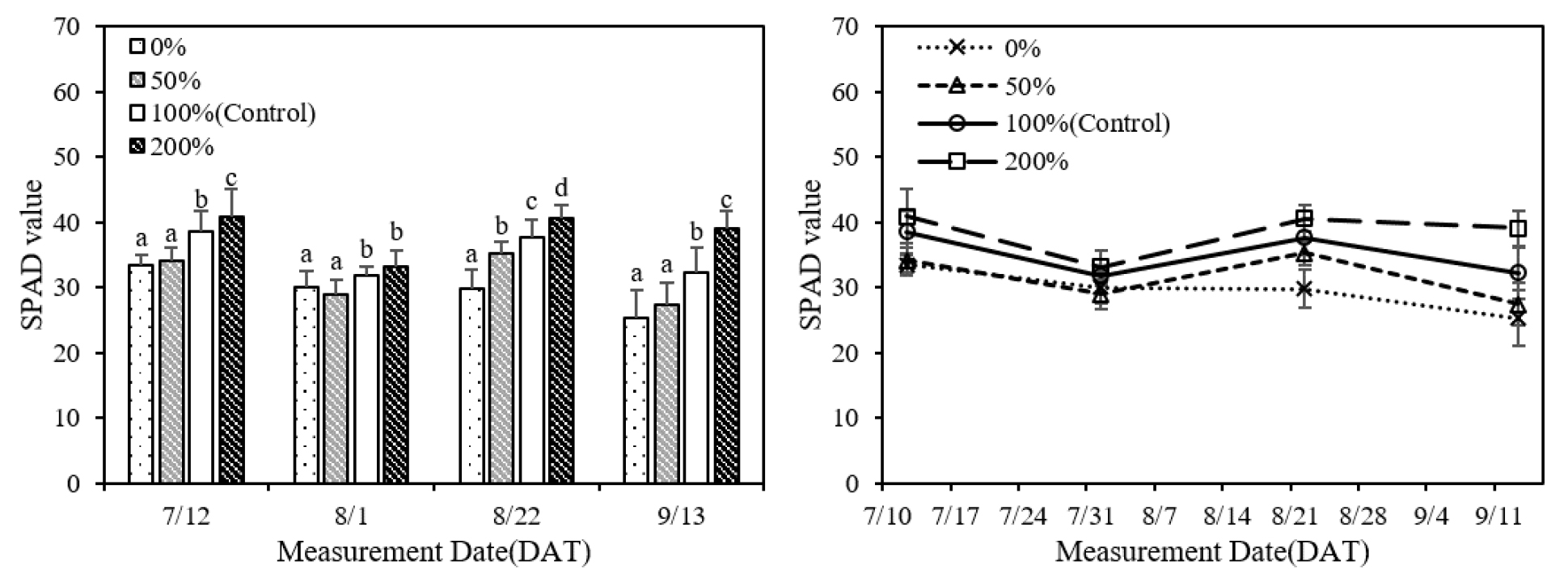
엽분광 데이터 분석
측정시기별로 질소시비량에 따른 분광 반사율을 분석한 결과, 시기마다 차이는 있었지만 질소시비량이 증가할수록 500~630 nm 및 680~730 nm 영역의 반사율이 감소하는 경향을 보였다(Fig. 5). 또한 분광 반사율과 1차 미분 값은 500~630 nm 및 710 nm 영역에서 질소시비량과 음의 상관관계를 나타냈다(Fig. 6). 광합성 관련 효소와 엽록소 농도가 증가하면 광 흡수율이 증가하고, 이에 따라 투과율과 반사율은 감소하지만 엽록소 농도가 특정 임계값에 도달하면 포화 효과(saturation effect)로 인해 청색과 적색 영역의 흡수 스펙트럼에서 반사율이 일정하게 유지된다(Gitelson & Merzlyak, 1996a). 본 연구에서도 430~480 nm 및 670~680 nm의 흡수 스펙트럼 영역에서 반사율의 차이가 작고, 질소시비량과의 상관관계가 낮았던 것은 포화 효과에 기인한 것으로 판단된다. 반면, 500~560 nm의 녹색 영역과 700~ 710 nm 영역의 반사율은 엽록소 a 농도에 민감하게 반응하여 엽록소 농도를 추정하는 지수 개발에 활용되었다(Gitelson et al., 1996b). 엽록소 a와 b는 주로 청색과 적색 영역의 빛을 흡수하지만, 식물체 내에는 많은 수의 엽록소 분자가 존재하기 때문에 더 넓은 범위의 광을 흡수 할 수 있다. 녹색 파장은 청색과 적색에 비해 흡수 효율이 낮지만, 엽록소에 의해 흡수되어 광합성에 사용된다(Hershey, 1995; Terashima et al., 2009; Virtanen et al., 2022). 또한 엽록소 농도가 증가하면 엽록소의 흡수 스펙트럼이 확장되어 흡수 스펙트럼 경계인 적색 가장자리 영역(red edge)의 광을 흡수한다(Banninger, 1991; Curran et al., 1991). 따라서 질소시비량 증가에 따른 엽록소 농도의 증가로 녹색 영역에 해당하는 500~600 nm 영역과 적색 가장자리 영역인 710 nm 영역의 반사율이 감소한 것으로 판단된다.
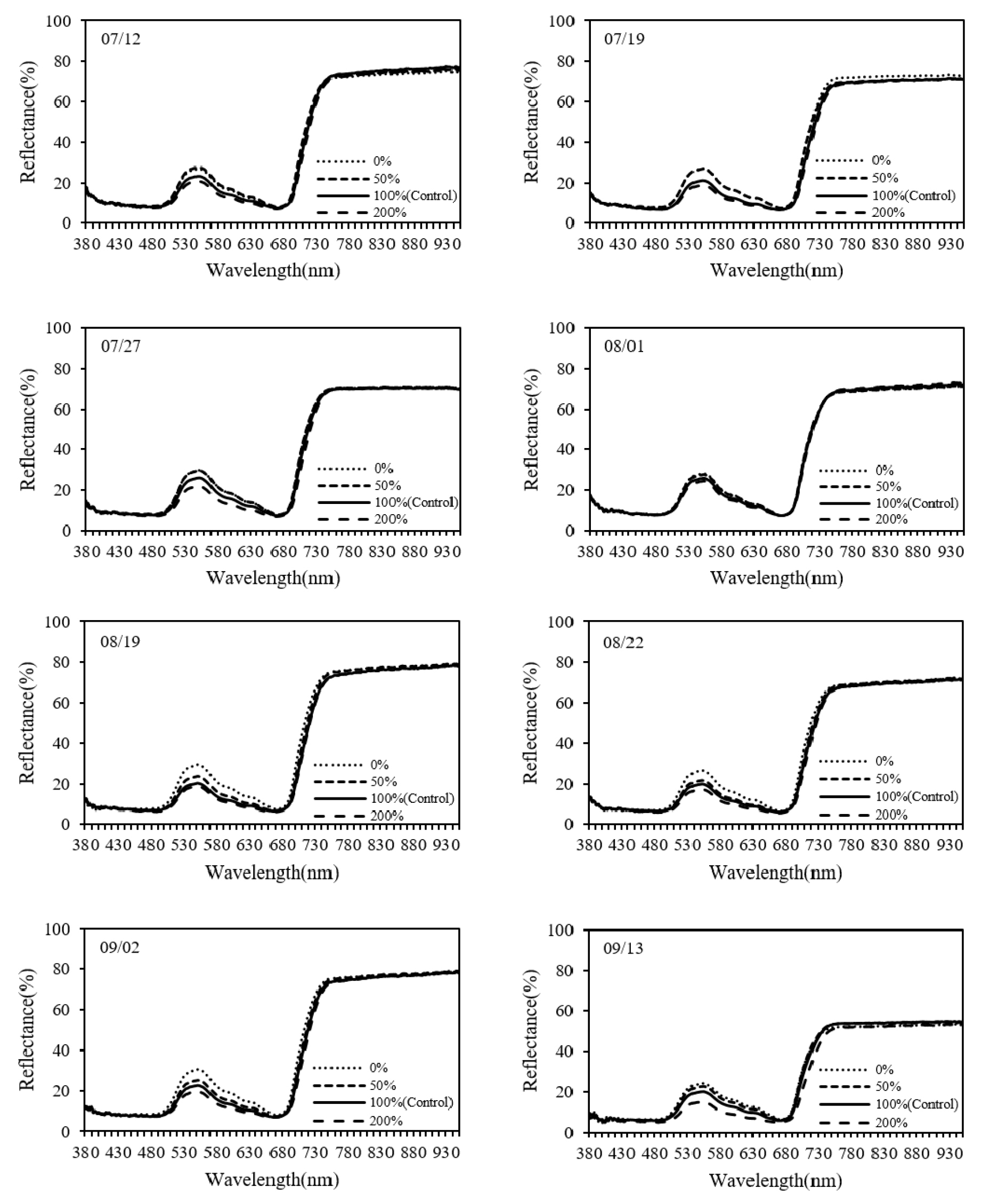
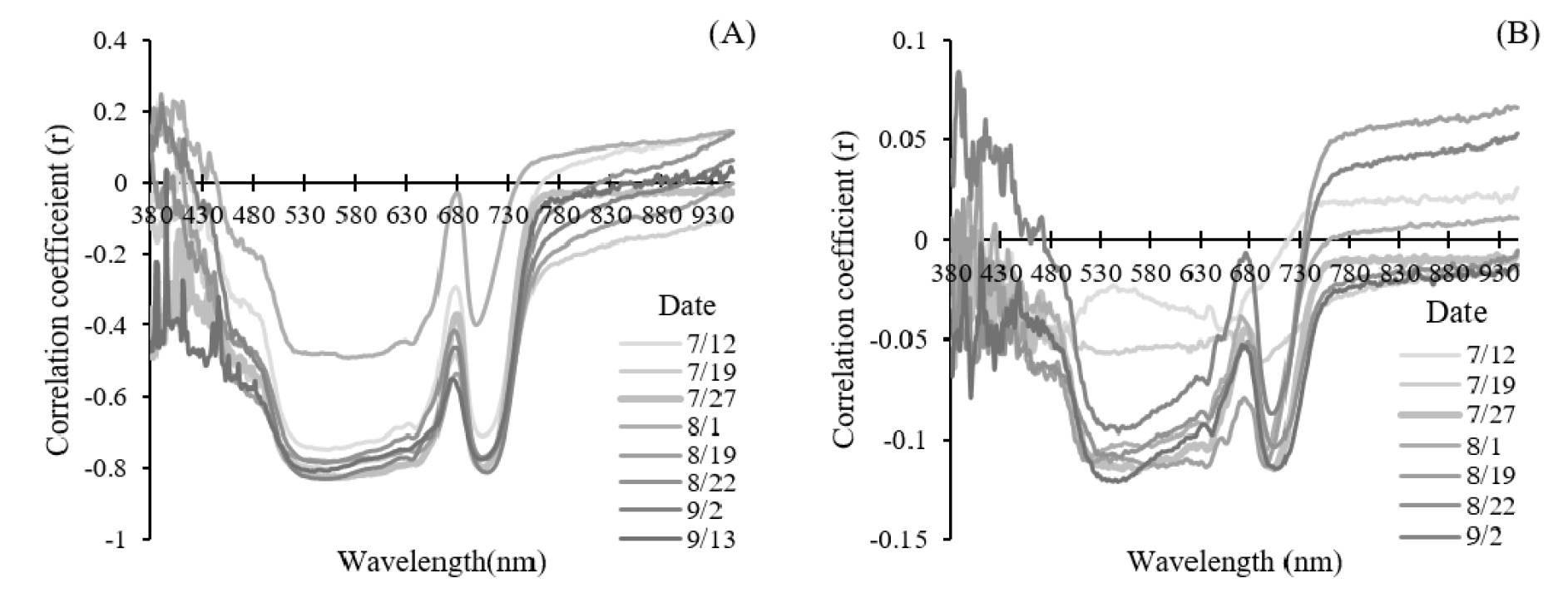
주요 파장 선택 및 모델 성능 평가
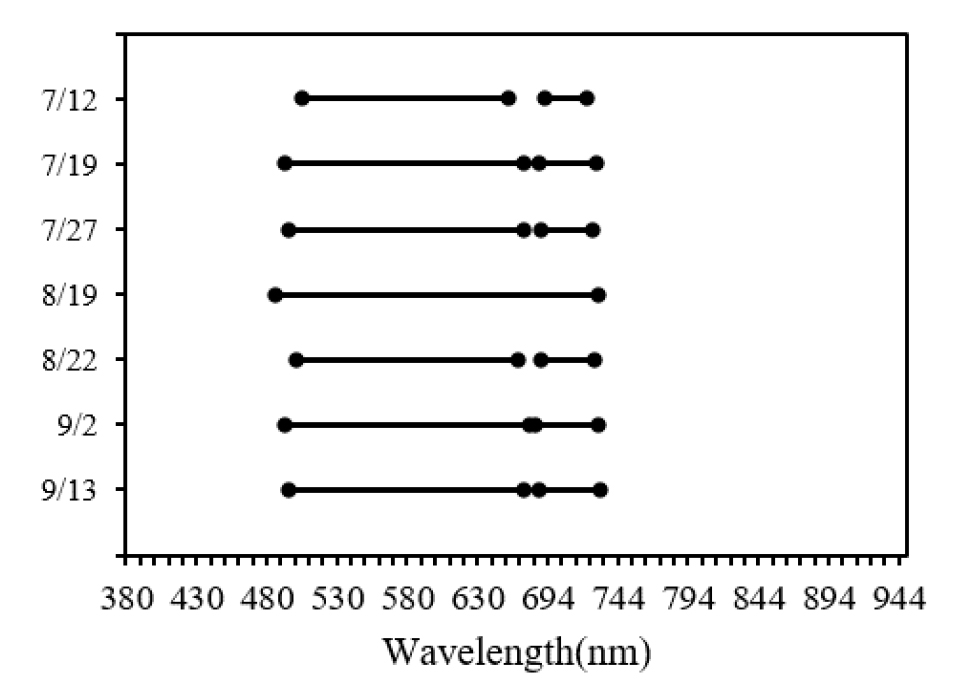
측정시기별로 처리 내 분산과 처리 간 분산을 비교한 결과 500~730 nm 영역을 1차적으로 추출하였으며 8월 1일 측정의 경우 반사율의 처리 간 분산이 처리 내 분산보다 크지 않았기 때문에 추출된 파장이 없었다(Fig. 7). 1차로 추출된 파장 영역 중에 상관성이 높은 파장을 그룹별로 분류한 뒤에 그룹별로 부분최소제곱법 분석을 통해 VIP 값을 계산하였고 측정시기별로 3~5개의 주요 파장을 선택하였다(Table 3).
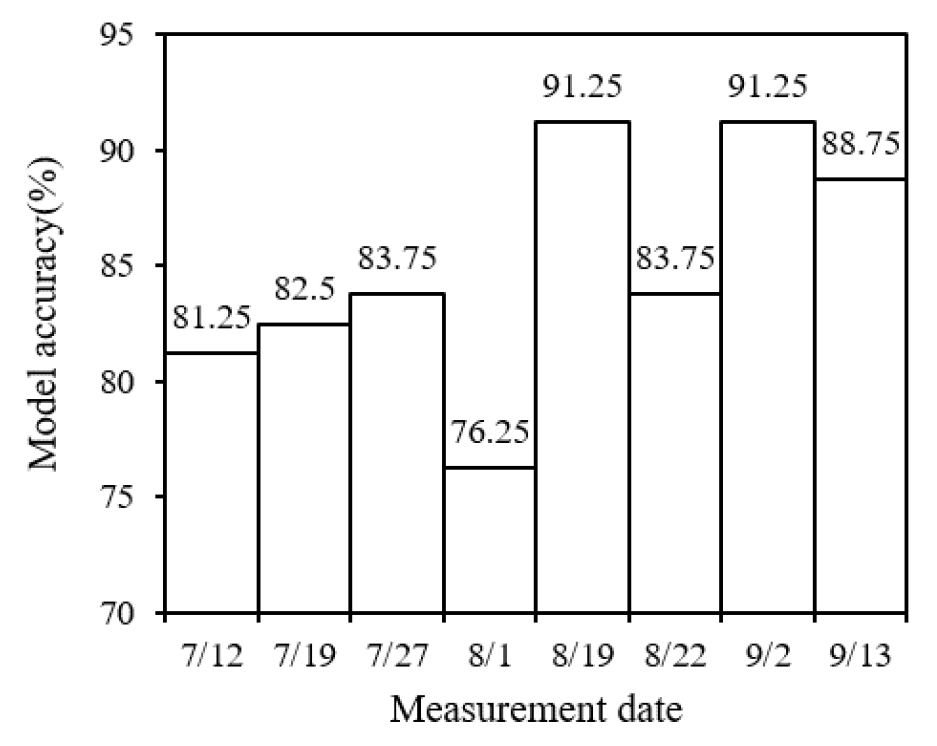
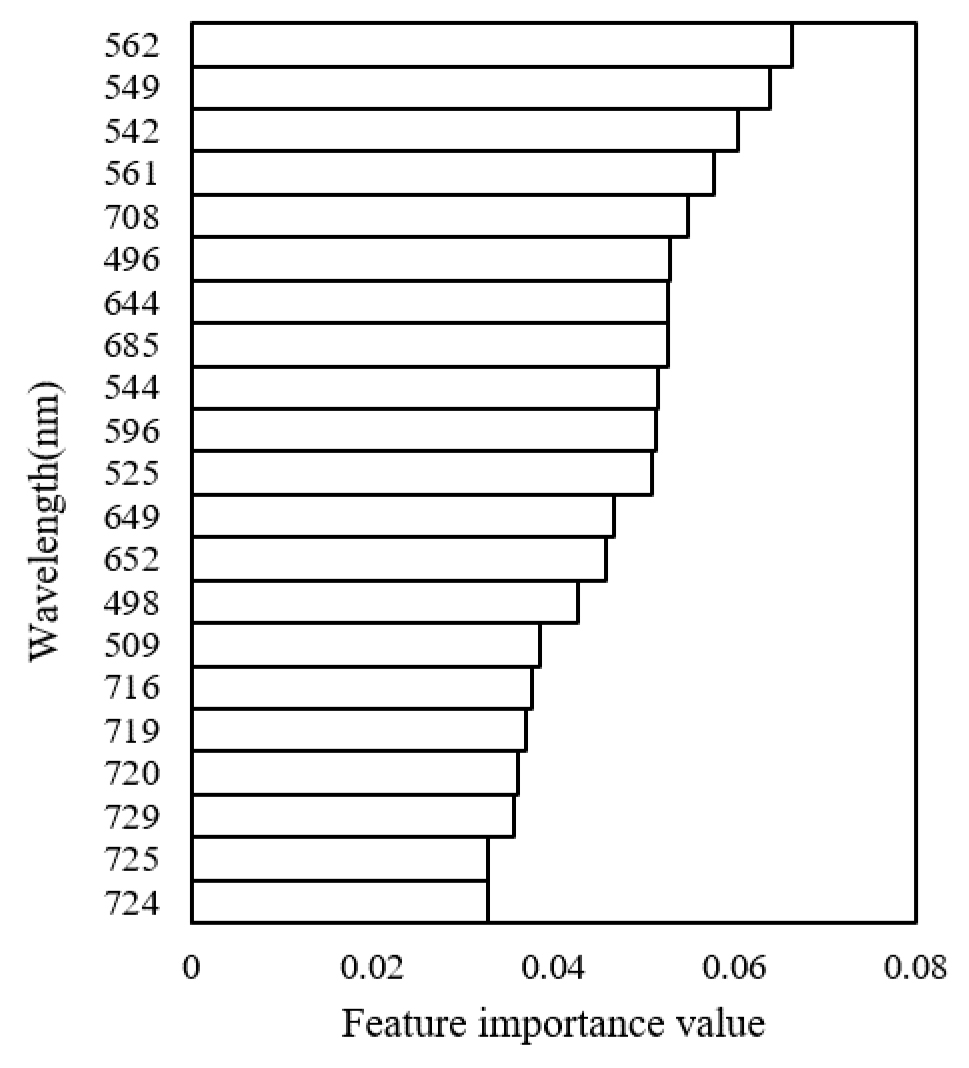
주요 파장을 입력 변수로 랜덤포레스트 분류 모델을 학습시킨 결과 평균 84%의 정확도를 확인하였으며 8월 19일과 9월 2일 모델의 분류 정확도가 91.25%로 가장 높았고 8월 1일이 76.25%로 가장 낮았다(Fig. 8). 또한 변수 중요도를 비교했을 때 550 nm 근처 파장의 중요도가 높게 평가되었고 720 nm 근처 파장의 중요도가 낮게 평가되었다(Fig. 9). 모델의 정확도는 변수 중요도가 높은 파장의 반사율이 처리 간 차이가 클수록 증가하였다. 반면에 8월 1일은 토양 질소 공급이 감소하고 질소 재분배로 잎의 엽록소 농도와 SPAD 값이 감소하면서 측정된 반사율의 처리 간 차이가 줄어들어 모델의 분류 정확도가 낮게 나타난 것으로 판단된다. 또한 생육 초중기보다 생육 후기에 모델의 정확도가 높게 나타났다. 이는 질소 결핍이 생육 후기에 잎의 노화를 유도하는 주요 환경 요인으로 작용하기 때문이며, 질소 시비 수준에 따른 잎의 엽록소 분해와 노화 속도의 차이가 생육 후기에 처리 간 반사율 차이를 증가시켜 모델의 분류 정확도가 높아진 것으로 보인다(Zakari et al., 2021). 또한 본 연구에서는 전체 측정시기의 데이터를 모델 학습에 사용하고, 각 측정시기별로 분류 정확도를 평가하였다. 이에 따라 추출된 파장이 포함되지 않은 8월 초순 데이터에서 분류 정확도가 낮게 나타난 것으로 판단된다.
결과적으로, 엽분광 반사율을 활용한 질소 시비 수준 분류 모델은 비료 시비 시기와 벼의 생육 단계에 따른 엽질소 함량 변화의 영향을 받는다. 따라서 보다 정확한 질소 시비 수준 분류를 위해, 측정시기는 시비한 비료가 고갈되기 전의 생육 초기, 그리고 질소 시비 수준에 따른 엽록소 분해와 노화 속도의 차이가 나타나는 생육 후기에 모델을 적용하는 것이 적절할 것으로 판단된다.
Table 3.
Wavelength selection by measurement period based on PLS analysis.
| Measurement date | 7/12 | 7/19 | 7/27 | 8/1 | 8/19 | 8/22 | 9/2 | 9/13 |
| Wavelength (nm) |
644 561 719 |
498 542 725 |
649 525 716 | - |
544 685 716 725 |
596 708 725 |
496 562 652 685 724 |
509 549 720 729 |
적 요
본 연구는 비파괴적인 방법으로 벼 포장의 질소 시비 수준을 평가하고자 엽분광계를 이용하여 벼 잎의 분광 반사율을 측정하고 랜덤포레스트 분류 모델을 학습시켜 분류 성능을 평가하였다. 질소시비량이 증가함에 따라 잎의 엽록소 농도가 증가하여 녹색 및 적색 가장자리 영역의 반사율이 감소하였다. 이를 기반으로 질소 시비 수준과 상관성이 높은 파장을 추출하여 랜덤포레스트 분류 모델의 입력 변수로 사용하였다. 분류 모델의 정확도는 반사율의 처리 간 차이가 클수록 증가하였으며, 생육 초중기에 비해 생육 후기에 더 높게 나타났다. 비료 시비 시기에 따라 식물이 이용할 수 있는 질소의 양이 달라지며, 출수와 등숙 과정에서 질소의 재분배와 잎의 노화로 인해 엽록소 농도가 변화하므로 측정시기에 따라 분류 정확도가 달라지는 것으로 판단된다. 따라서 질소시비량에 따른 엽록소 농도의 변화가 크게 나타나는 생육 후기에 분광 반사율을 이용하여 질소 시비 수준을 평가할 수 있을 것으로 판단된다. 향후, 모델의 정확도를 높이기 위해 특정 생육시기마다 처리별로 충분한 분광 반사율 데이터를 수집한다면, 각 생육시기별로 질소 시비 수준을 더 정확하게 분류할 수 있을 것이다.
