

서 론
녹두[Vigna radiata (L.) R. Wilczek]는 콩과(Fabaceae) 비냐속(Vigna)에 속하는 일년생 작물로, 인도 인근에서 기원하여 중·동·남아시아 지역을 중심으로 재배된다. 녹두는 단백질, 철분, 미네랄 등 다양한 영양성분이 풍부하여 영양학적으로 중요한 작물로 평가된다(Dahiya et al., 2015; Yi-Shen et al., 2018; Hou et al., 2019). 또한, 생육기간이 짧고 기후 적응성이 높아 넓은 범위의 위도에서 재배가 가능하며, 여러 작부체계에서 활용된다(Sandhu & Singh, 2021; Dhaliwal et al., 2023). 그러나 재배화 이후 녹두 품종들은 유전적 다양성이 야생종에 비해 약 30% 수준까지 감소한 것으로 보고되었으며(Ha et al., 2021), 기존 연구에서도 유전적 다양성 확보의 한계가 지적되어 왔다(Kim et al., 2015; Gayacharan et al., 2020). 이러한 좁은 유전적 기반은 기후변화로 인한 환경 스트레스 대응력 약화로 이어질 수 있다(Van Haeften et al., 2021). 최근 기후변화로 인해 주요 10대 작물의 전 세계 생산량이 약 1% 감소한 것으로 보고되면서, 기후변화 적응형 품종 개발의 필요성이 더욱 강조되고 있다(Rivero et al., 2022). 이러한 중요성을 고려할 때, 유용 형질을 가진 품종 육성을 위해서는 충분한 유전적 변이와 다양한 형질 평가가 가능한 육종 소재의 확보가 필수적이다(Azam et al., 2023; Irfan et al., 2025).
그러나 전 세계 약 1,750개의 유전자원 은행에 약 740만 점의 자원이 보존되어 있음에도, 실제 육종에 활용되는 비율은 약 2%에 불과한 것으로 보고되었다(Mondal et al., 2023). 이러한 낮은 활용도는 보유 자원에 대한 정밀한 표현형 평가가 충분히 이루어지지 않았기 때문이며, 이는 현재 육종의 주요 한계 중 하나임을 시사한다. 따라서 유전자원의 활용도를 높이기 위해서는 육종 목표 환경에서 정밀한 표현형 평가와 다변량 분석을 통한 변이 구조 파악이 필요하다(Sandhu & Singh, 2021). 한편, 지금까지 수행된 녹두 유전자원 표현형 평가 연구들은 주로 인도, 중국, 인도네시아 등 일부 지역에 편중되어 있으며, 분석규모 또한 50-180점 수준으로 제한적이었다(Rasal & Parhe, 2017; Wang et al., 2018; Jadhav et al., 2021; Manjunatha et al., 2023). 국내에서는 한국, 인도, 중국 3개국 323점을 평가한 보고(Desta et al., 2024)가 있으나, 보다 다양한 기원을 포괄한 양적형질의 평가와 변이 구조 분석의 확장이 필요하다.
이에 본 연구에서는 국내외 12개국에서 수집된 477점의 녹두 유전자원을 대상으로 생육 및 수량 관련 양적형질을 평가하고, 상관분석, 주성분분석(PCA), 군집분석 등 다변량 통계분석법을 통해 양적형질의 변이 구조와 형질 간 관계를 분석하였다. 이를 통해 국내 재배환경에 적합한 품종 개발을 위한 기초자료를 제공하고자 한다.
재료 및 방법
실험재료
본 연구에서는 녹두 477점을 농촌진흥청 농업유전자원센터, 세계채소센터(WorldVeg)에서 분양 받아 이용하였다. 유전자원은 부여된 등록번호(IT, VI 등)로 표기하였고, 수집국가를 표기하였다(Supplementary Table 1). 시험에서 사용한 477점의 녹두 유전자원은 기원지인 인도를 중심으로 파키스탄, 아프가니스탄 등 12개국의 다양한 국외 유전자원 214점과 한국 유전자원 263점이었다.
재배 방법 및 농업형질 조사
본 연구는 농촌진흥청 국립식량과학원 본원(전북 완주군) 포장에서 수행하였으며, 2024년 6월 21일 파종하였다. 각 자원은 자원당 7주, 1주 1본으로 파종하였으며, 조간 150 cm × 주간 15 cm로 재배하였다. 이 재식거리는 유전자원 간 교잡 방지와 개체 간 간섭을 최소화하여 형질 평가의 정확성을 높이기 위해 설정하였다. 농업형질 조사는 농촌진흥청 농업과학기술 조사 분석기준(RDA, 2012)과 World Vegetable Center(WorldVeg) 조사 분석기준 일부를 수정하여 이용하였다(Table 1). 조사 대상 형질은 모두 양적형질로서, 개화일수(days to flowering, DTF), 성숙일수(days to maturity, DTM), 개화 후 성숙까지의 일수(days from flowering to maturity, FTM), 협장(pod length, PL), 협당립수(number of seeds per pod, SPP), 백립중(one-hundred seeds weight, HSW)을 포함하였다.
Table 1.
Description of six quantitative traits evaluated in mungbean germplasm accessions.
통계분석
본 연구의 통계분석은 R studio software (www.r-project. org, v.4.4.2., Vienna, Austria)를 이용하였다. 양적형질 간 상관관계분석은 Pearson 상관계수로 유의성을 확인하였고, 녹두 유전자원 집단의 특성과 분포를 설명하기 위하여 수치간 표준화를 통하여 주성분 분석을 수행하였다. 군집분석은 유클리드(Euclidiean distance) 거리 계산법에 기반하여 최장 연결법(complete linkage method)을 적용하여 군집화 하였다.
결과 및 고찰
녹두 유전자원의 생육특성 변이
녹두 유전자원 477점에 대해 총 6가지 양적형질을 조사한 결과, 한국 자원과 국외 자원 간 모든 형질에서 유의한 차이(p<0.01)가 나타났다. 각 형질에 대한 최소값, 최대값, 평균, 표준편차 및 변이계수는 Table 2에 제시하였다. 전체 자원에서 DTF는 36-92일의 범위를 보였으며, 이는 기존 보고보다 더 넓은 분포를 보였다. 이는 기원지인 인도를 포함한 12개국의 다양한 기원의 유전자원을 포함하여 유전적 변이의 폭이 상대적으로 컸기 때문으로 판단된다(Zhao et al., 2021; Manjunatha et al., 2023; Desta et al., 2024). 기원에 따른 비교 결과, DTF는 한국 자원(평균 46.7일)이 국외 자원(64.5일)보다 유의하게 짧았고, 개화 이후 성숙까지의 FTM은 각각 19.7일과 23.4일, DTM은 65.4일과 86.8일로 나타나, 한국 자원이 전반적으로 짧은 생육 소요일을 보였다. 이러한 결과는 한국 자원이 생식생장기로의 전환이 상대적으로 빠른 조숙형 생육 특성임을 보여준다. 한편, 국외 자원의 상당수가 저위도(중∙동∙남아시아) 기원임을 고려할 때, 상대적 장일 조건인 한국 재배환경에서 단일성(short-day)작물인 녹두의 광주기 반응에 의해 개화와 성숙이 지연된 결과로 해석된다(Vas Aggarwal & Poehlman, 1977; Amkul et al., 2024).
Table 2.
Basic statistics for quantitative traits of 477 mungbean accessions of Korea and foreign origins.
생육기간과는 달리 생산성과 관련된 형질에서는 반대의 경향이 관찰되었다. PL은 5.2-13.2 cm의 범위에서 분포하였으며, 한국 자원의 평균은 9.09 cm로 국외 자원(7.76 cm)보다 유의하게 길었다(p<0.01). SPP와 HSW 또한 각각 13.49립 및 4.56 g으로, 국외 자원(12.75립, 3.92 g)에 비해 유의하게 높은 값을 보였다. 분포 특성에서도 국외 자원의 분포는 첨도(kurtosis)가 높아 중심값 부근에 집중된 반면, 한국 자원은 상대적으로 첨도가 낮고 분산이 크기 때문에 다양한 표현형 변이를 포함하는 것으로 나타났다.
종합적으로 보았을 때, 한국은 위도 33°-42°에 위치한 사계절성 기후권에 속하며, 녹두의 재배기는 여름철 고온에서 시작해 가을로 접어들며 급격히 저온화되는 짧은 생육기간 특성을 나타낸다. 특히 국내 파종 적기인 6월 하순은 하지 직후로, 생육초기에 상대적으로 장일을 거친 뒤 점차 단일로 전환되는 광주기 조건에서 생육한다. 이러한 환경은 생식생장으로의 전환을 빠르게 유도하는 생리적 적응이 이루어졌을 가능성이 크며, 본 연구에서 한국 자원에서 조숙성이 상대적으로 빈번하게 관찰된 결과와 부합한다. 반면, 국외 자원의 상당수는 저위도 지역의 단일 환경에 적응한 자원으로, 한국의 상대적 장일조건에서는 개화 유도가 지연되고 결과적으로 성숙이 늦어져 생산성이 감소하는 경향을 보였다. 실제로 단일성 콩과 작물에서 장일 조건이 수확지수를 감소시킬 수 있음이 보고된 바 있으며(Chauhan & Williams, 2018), 본 연구에서도 동일한 경향이 나타났다.
상관관계 분석
녹두 유전자원의 양적형질에 대한 상관관계를 분석한 결과, 대부분의 형질 간 조합에서 p<0.01 수준의 유의한 상관이 확인되었다(Fig. 1). 생육기간 관련 형질인 DTF, FTM, DTM은 서로 양의 상관을 보였으며, 특히 DTM과 DTF 간에는 매우 높은 양의 상관(r = 0.96)이 나타났다. 이는 DTM가 DTF에 크게 좌우됨을 의미한다. 한편, FTM은 DTF 및 DTM과 각각 r = 0.46, r = 0.69로 통계적으로 유의하였으나 상대적으로 낮은 상관을 보여, 개화 후 성숙 기간이 개화소요일수와 완전히 비례하지 않음을 나타낸다. 이는 등숙기의 환경 요인에 의해 FTM이 일부 자원에서는 독립적으로 조절된 결과로 해석된다.
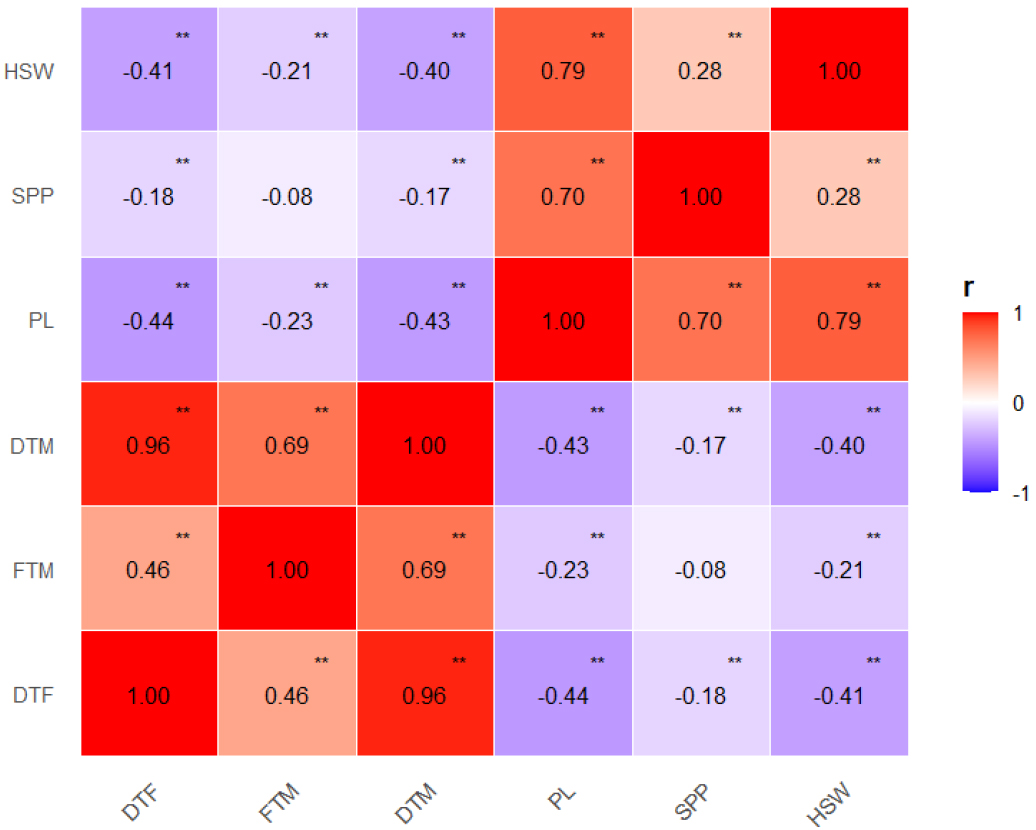
Fig. 1.
Pearson correlation matrix for six quantitative traits of 477 mungbean accessions. Positive and negative correlations are indicated by red and blue colors, respectively, with darker shades representing stronger associations. DTF, days to flowering; FTM, days from flowering to maturity; DTM, days to maturity; PL, pod length; SPP, number of seed per pod; HSW, 100-seed weight. *p < 0.05, **p < 0.01.
실제로 한국의 녹두 성숙기는 8월에서 10월 말에 이르며, 이 시기의 평균 기온은 최적 생육온도(28-30℃)에 비해 낮다. 등숙기의 저온은 광합성 및 동화산물 전류를 저하시켜 등숙을 지연시키므로, 늦게 개화한 유전자원일수록 성숙이 지연되는 경향을 보였을 가능성이 크다(Manasa et al., 2023). 따라서 한국의 생육 후반 환경 조건으로 인해 FTM은 DTM과 완전한 선형성을 보이지 않은 것으로 해석된다.
생산성과 관련된 형질인 PL은 SPP, HSW과 서로 강한 양의 상관을 보였다. 반면 SPP와 HSW간의 상관은 상대적으로 낮았다(r = 0.28). 즉, 협의 길이가 길어지면 보통 종자 수와 종자비대가 함께 증가하지만, 협 내 종자 수와 종자 비대 간에는 일정 부분 독립적인 경향이 있음을 시사한다. 반면, 생육기간 관련 형질(DTF, FTM, DTM)과 생산성 관련 형질(PL, SPP, HSW)간에는 모두 음의 상관이 나타났다. 이는 기존의 보고와 일치되는 결과였다(Azam et al., 2023). 즉, 개화 및 성숙이 늦은 자원일수록 협의 발달과 종실의 비대가 제한되어 상대적으로 낮은 생산성을 보이는 경향이 있었다. 한국 자원과 국외 자원을 구분하여 상관관계 분석을 한 결과, 전반적인 상관 구조는 유사하였으나, PL과 SPP의 상관관계는 국외 자원(r = 0.46)에 비해 한국 자원(r = 0.75)에서 더 높게 나타났다(Supplementary Fig. 1). 이러한 차이는 자원의 기원에 따라 생산성 관련 형질 간 상호작용이 서로 다르게 형성되었음을 시사한다.
주성분 분석
여러 형질의 다차원적 변이를 대표적인 주성분(principal components)로 축약하여 유전자원 간 변이 패턴을 시각적으로 비교하고자 주성분 분석(PCA)을 수행하였다(Venujayakanth et al., 2017). 분석 결과, 고유값(eigenvalue)이 1이상인 주 성분은 총 두 개이며, 제 1주성분(PC1)과 제 2주성분(PC2)은 전체 변이의 53.87%와 24.35%를 설명하여, 전체 변이의 약 78.22%를 설명하였다(Table 3).
Table 3.
Eigenvalues, proportion of variance, and principal component loadings of quantitative traits for mungbean germplasm accessions.
PC1은 DTF(+0.462)와 DTM(+0.484)가 양의 방향으로, PL(-0.448)과 HSW(-0.399)가 음의 방향으로 높게 기여하였다. 이는 생육기간 관련 형질과 수량 관련 형질이 서로 반대 방향의 변이를 보이며 자원 간 분포 차이를 형성하고 있음을 의미한다. PC2는 FTM(-0.420), PL(-0.457), SPP(-0.532)의 기여도가 컸으며, 등숙기간과 수량 관련 형질의 변이를 함께 반영하는 축으로 나타났다. 이러한 결과는 자원 간 변이가 개화 이후의 생육기간과 수량 관련 형질에 의해 추가적으로 구분됨을 시사한다.
두 주성분을 기준으로 각 자원의 기원을 구분하여 시각화한 결과, 국외 자원은 PC1의 양의 방향, 한국 자원은 PC1의 음의 방향으로 중심이 치우쳐 부분 분리를 나타냈다(Fig. 2). 이는 기술통계에서 분석된 바와 같이, 국외 자원이 상대적으로 늦게 개화하고 협 및 종실 발달이 상대적으로 약한 자원이 많으며, 반대로 한국 자원은 개화가 비교적 빠르고 협장 및 백립중이 큰 자원이 상대적으로 많다는 경향과 일치한다. 기원지 별 95% 신뢰 타원 영역은 일부 중첩되었으나, 중심과 분포 방향이 상이하여 기원에 따른 표현형 변이 구조가 부분적으로 구분됨을 나타낸다. 또한 두 집단 모두 타원의 면적이 넓어, 각 집단 내에서도 폭넓은 변이가 존재하는 것으로 보이며, 이는 다양한 동일한 기원 내에서도 다양한 소재 탐색 가능성이 높음을 의미한다. 이러한 결과는 PCA를 통해 각 자원이 어떤 형질군이 각 자원의 분산을 주도하는지 명확히 제시하며, 유전자원의 기원이 양적형질의 변이 분포 양상을 설명하는 중요한 요인이 될 수 있음을 시사한다.
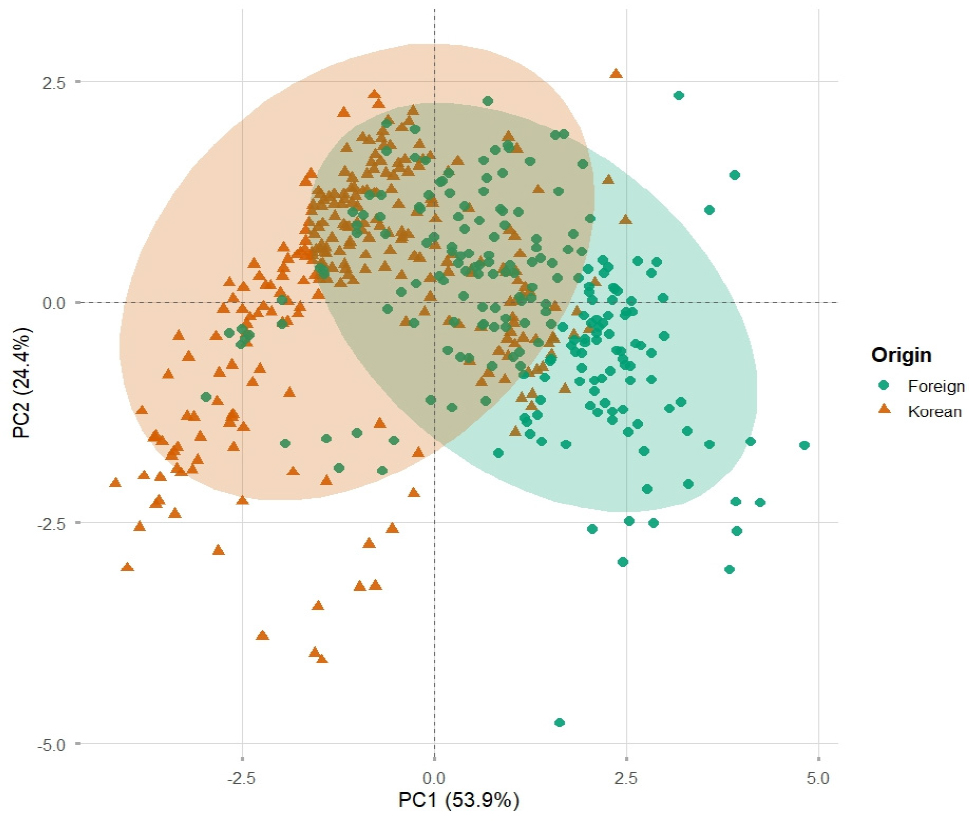
Fig. 2.
Principal component analysis based on six quantitative traits of 477 mungbean accessions. The first and second principal components explained 53.87% and 24.35% of the total variance, respectively. Each point represents an individual accession, colored according to its origin (green for foreign and orange for Korean accessions).
군집분석
Elbow method를 적용한 결과, 군집 내 제곱합(Total within-cluster sum of squares, WSS) 감소폭이 k = 3 이후부터 완만해져 최적 군집수를 3으로 설정하였다(Supplementary Fig. 2). Euclidean 거리를 이용한 계층적 군집분석에서도 동일하게 세 개의 군집이 확인되어, 서로 다른 접근법에서도 일관된 분화 구조가 확인되었다(Supplementary Fig. 3). 이는 군집 분석 결과의 높은 신뢰성을 뒷받침한다.
K-means 군집분석 결과, 녹두 유전자원 477은 뚜렷한 세 개의 군집으로 구분되었다(Fig. 3). 군집I은 전체의 38.99% (186점)를 차지하였으며 군집 내 국외 자원 비율이 74.73%로 높았다. 군집II는 전체의 42.35%(202점)로 가장 큰 군집이었으며, 한국 유전자원이 70.3%를 차지하였다. 군집III은 전체의 18.66%(89점)으로 가장 작은 군집이었으나, 한국 자원 비율이 83.15%로 가장 높게 나타났다(Supplementary Table 2).
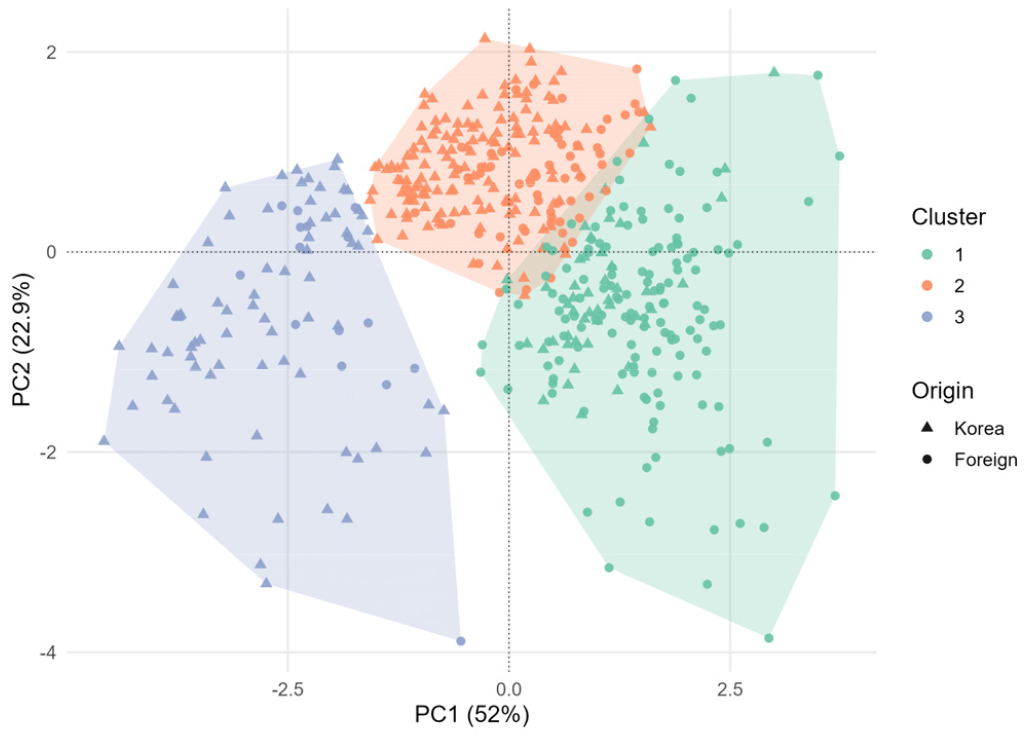
Fig. 3.
K-means clustering of 477 mungbean accessions based on quantitative traits. Each cluster is represented by a different color: green (I), orange (II), and blue (III). The symbols indicate the origin of accessions, with triangles representing Korean accessions and circles representing foreign accessions.
군집별 형질 평균값 비교 결과(Table 4), 군집I은 DTF, FTM, DTM이 각각 69.13일, 25.59일, 93.72일로 가장 길어 세 군집 중 가장 늦게 성숙하는 집단이었다. 수량 관련 형질인 PL, SPP, HSW는 가장 낮아 생육 기간은 길고 수량성은 낮은 경향을 보였다. 군집II는 군집I에 비해 DTF, FTM, DTM이 모두 짧았으며, PL과 HSW는 군집I과 군집III의 중간 수준, SPP는 군집I과 유사한 수준으로 나타났다. 군집III은 DTF, FTM, DTM이 각각 45.40일, 19.63일, 64.03일로 나타났으며, 이는 군집II와 유사한 수준이었다. 반면 군집 I와 비교하면 DTF, FTM, DTM이 각각 23.73일, 5.96일, 29.69일 더 짧아 생육 기간에서 현저한 차이를 보였다. 또한 군집III은 수량 관련 형질인 PL, SPP, HSW에서는 세 군집 중 가장 높은 값(PL: 11.18 cm, SPP: 14.73개, HSW 5.90 g)을 나타내었다. 군집 I과 군집II에 비해 PL은 2.98-3.67 cm, SPP는 1.84-2.03개, HSW는 1.81-2.21 g 더 높아 조숙성과 다수성이 가장 두드러지는 집단으로 평가되었다.
Table 4.
Mean value of quantitative traits among three k-means clusters.
녹두는 일반적으로 생육 최적온도인 28-30℃에서 안정적으로 등숙한다. 그러나 한국 환경에서는 수확기가 늦어질수록 기온이 최적온도보다 낮아져, 병해 발생과 종실의 품질 저하로 인해 생산 안정성이 떨어지는 경향이 있다. 이러한 생리적 특성을 고려할 때, 국내 녹두 품종 개발에 있어 개화가 늦고 수량성이 비교적 낮은 군집 I의 자원은 활용도가 낮은 반면, 조숙하며 수량성이 높은 군집III 자원은 국내 환경에 적합한 품종 개발 소재로 활용할 가치가 높을 것으로 판단된다.
적 요
1.본 연구는 국내외에서 수집된 녹두[Vigna radiata (L.) Wilczek] 477점의 유전자원을 대상으로 생육 및 수량 관련 양적형질을 평가하여 변이 구조와 집단 간 특성을 분석하였다.
2.한국 자원은 국외 자원보다 생육기간이 짧아 조숙형 특성을 보였으나, 협장, 협당립수, 백립중이 높아 한정된 생육기간 내에서도 효율적인 협 발달과 종실비대가 이루어졌다.
3.주성분 분석 결과, 전체 변이의 약 78.22%가 두 개의 주성분(PC1, PC2)에 의해 설명되었으며, 한국 자원과 국외 자원은 개화가 늦고 생산성 관련 형질이 낮은 특성이 반영된 PC1 축을 따라 부분적으로 분리되는 경향을 보였다.
5.군집분석에서 3개의 군집이 확인되었으며, 군집III은 조숙형이면서 다수성의 특성이 두드러져 국내 재배환경에 적합한 육종 소재로 평가되었다.
6.본 연구 결과는 녹두 유전자원의 생리·형태적 다양성에 대한 이해를 확장하고, 국내 환경에 적합한 품종 개발을 위한 기초자료로 활용될 수 있을 것이다.
