

서 론
재료 및 방법
실험재료 및 처리내용
시험구 배치 및 조사방법
기상자료
통계분석
결과 및 고찰
콩 재배 기간의 기상변화
파종 시기별 출현율 및 출현소요일수
파종시기별 온도와 일장 변화에 따른 생육반응
생육, 수량구성요소 및 수량
적 요
서 론
콩(Glycine max Merr.)은 콩과에 속하는 전형적인 단일성 작물로 풍부한 단백질 및 식물성 오일 공급원으로 전 세계에서 재배되고 있다(Medic et al., 2014). 국내 콩 재배면적은 2023년 6.8만 ha이고, 총 생산량 14.1만 톤으로 2018년도의 6.3만 ha, 10.6만 톤에 비해 재배면적과 생산량이 증가하고 있다(KOSIS, 2023). 최근 정부의 전략 작물 직불제 시행으로 벼 대신 논에 콩을 재배하거나, 특히 남부지역의 경우에 온난한 기후 여건과 더불어 소득향상을 위해 봄배추, 봄감자, 옥수수 등 다양한 작물과의 작부체계에 콩을 도입하고 있다. 우리나라에서 콩은 지역에 따라 다소 차이가 있지만 일반적으로 6월 초․중순~하순경에 파종하면 7월 하순~8월 상순에 개화하고, 10월 하순경에 수확하는 형태가 대부분이다. 그러나 최근 기후변화로 콩 파종시기인 5~6월의 상습적인 가뭄이나 7~8월의 기습적인 폭염, 폭우 등으로 인한 출현율 저하 및 개화 지연과 같은 피해가 발생하고 있다. 실제 미국 콩 재배지역에서 30년간(1967~2006년) 온도변화에 따른 수량을 조사한 결과 온도가 1℃~0.8℃ 상승하면 수량이 약 1.6%~2.4%정도 감소한다고 하였다(Kucharik & Serbin, 2008; Hatfield et al., 2011). 이러한 기후변화에 대응하기 위한 가장 용이한 방안 중 하나가 파종시기의 이동이다(Kim et al., 2013). 파종시기를 이동하게 되면 콩의 생육 환경이 변하게 되는데, 이때 온도와 일장이 파종시기에 따라 달라지게 된다. 콩에서 일반적으로 장일과 고온은 콩의 영양생장을 촉진시키는 반면에 단일과 저온은 생식생장을 촉진시킨다(Wang et al., 2022). 콩의 온도는 일반적으로 15℃~25℃ 사이에서는 온도가 올라갈수록 개화가 단축되고 평균온도가 20℃ 이하이거나 30℃ 이상이 되면 개화가 지연된다(Gibson & Mullen, 1996). 일장은 개화 및 성숙과정에도 영향을 미쳐 조기 파종을 하면 마디수, 초장이 증대되고, 조기 개화를 하며, R1~R5 단계가 길어져 수량 증대 효과가 있다(Sinegovskaya & Levina, 2021; Schoving et al., 2020). 하지만 조기 파종시 적절한 재배관리를 하지 않으면 온도가 높고, 일장이 길어 영양생장 기간이 늘어나서 도복이 발생하고 불량한 착협을 유도하여 오히려 수량이 감소하기도 한다. 반면 파종시기가 적기보다 늦어지면 생육후기 온도가 낮아지고 일장이 짧아지기 때문에 수량이 감소하지만 초장 단축이나 고온 회피 등으로 재배관리가 쉬운 장점도 있어 기후변화에 따른 콩의 파종시기별 생육 및 수량특성 구명이 필요하다(Kantolic et al., 2013; Nico et al., 2015).
따라서 본 연구는 최근 기후변화에 대응하여 남부지역에서 적기파종 이후 파종 시기를 달리하였을 때 온도와 일장이 개화 및 등숙일수와의 관계를 구명하고 더불어 파종시기가 수량에 미치는 영향을 알아보고자 수행하였다.
재료 및 방법
실험재료 및 처리내용
본 시험은 2023년도 한반도 남부지역 경남 진주(35°11′34″N / 128°5′5″E)에서 수행하였다. 공시 품종은 조생종 ‘새올’, 중생종 ‘선유 2호’, 중만생종인 ‘대원’ 등 총 3품종을 이용하였다. 파종시기는 6월 20일부터 약 10일의 간격을 두고 7월 1일, 7월 10일, 7월 20일, 7월 30일, 8월 10일 등 총 6회를 순차적으로 파종하였다. 시비는 전량 기비로(시비량 : N ‒ P ‒ K = 3 ‒ 3 ‒ 3.4 kg/10a) 시비하고 재식밀도는 밀식에 의한 영향을 최소화 하기 위해 80 cm × 20 cm로 1주 2본을 파종하였으며, 흑색 비닐을 이용한 피복재배를 하였다. 기타재배법은 농촌진흥청 표준재배법에 준하였다.
시험구 배치 및 조사방법
시험구 배치는 조생종, 중생종, 만생종 등 3품종을 난괴법 3반복(30주 1반복)으로 배치하였다. 출현율은 출현수/파종립수를 백분율로 나타내었고, 출현 소요일수는 50% 출현한 날짜를 소요일수로 나타내었으며, 출현율은 파종 후 15일 동안 출현한 개체 수를 조사하였다. 개화시는 첫 꽃이 개화한 날, 개화기는 50% 이상 개화한 날로 조사하였다. 수확은 처리별 수확기(R8; 95%꼬투리가 숙색을 나타낼 때)에 수확하였으나 7월 30일, 8월 10일 파종은 저온으로 더 이상 등숙이 진행되지 않아 몇몇 조사주 수(저온에 대한 등숙 현황)를 제외하고 11월 11일에 수확하였다. 수량은 샘플 개체에 대해 조사된 수량구성요소의 산출식[수량 = (주수 × 협수 × 협당립수 × 백립중) / 100]을 통해 10a당 수량을 구하였다.
기상자료
기상청의 기상자료 개방포털(KMA, 2023)에서 진주시의 2023년 기상자료를 진주지역의 10년 평년 온도와 강수량(2013-2022)과 비교했다.
통계분석
모든 결과는 10주를 1반복으로 하여 3 반복 측정하였으며, 평균과 표준편차 또는 표준오차로 나타내었다. 통계분석 및 다중회귀 분석은 SPSS program (SPSS version 21, SPSS Inc., Chicago, IL, USA) 프로그램을 이용하여 일원배치 분산분석(one way ANOVA)을 실시한 후, Duncan’s Multiple Range Test (DMRT)을 통해 5%, 1%, 0.1% 수준에서 통계학적 유의성을 검정하였다. 다중회귀 분석은 콩 파종시기에 따라 파종~개화기, 개화기~수확기까지의 조사된 생육일수를 종속변수로 이용하였고, 독립변수는 파종시기별 소요일수에 해당하는 누적일장과 적산온도를 이용하였다. 상관 관계분석은 R (R studio server pro 4.2.2)의 ‘corrplot’ 이용하여 분석하였고, 상관관계와 다중회귀분석은 모든 생태형을 합산하여 분석하였다.
결과 및 고찰
콩 재배 기간의 기상변화
진주 지역의 연구 기간(2023년) 동안의 기상 현황을 보면, 6월부터 점차 기온이 상승하여 7월은 26.6℃, 8월은 28.0℃로 1년 중 가장 더웠으며, 평년의 7월 25.9℃, 8월 26.9보다 각각 0.7℃, 1.1℃ 높았다(Fig. 1). 8월 이후 기온은 점차 하강하여 11월에는 9.6℃까지 떨어졌으며 평년의 9.0℃ 대비 0.6℃ 높아 전체적으로 평년 대비 기온이 높았다. 시험기간 동안 총 강수량은 1,462.5 mm로 평년의 1,058.9 mm보다 403.6 mm 많았다. 특히 개화기인 8월, 9월에 각각 439.5 mm, 200.5 mm로 평년의 251.4 mm, 180.3 mm 비해 많았으며 등숙기인 10월은 8.5 mm로 평년의 105.0 mm에 비하여 강수량이 매우 적었다(Fig. 1).

파종 시기별 출현율 및 출현소요일수
조생종에서 출현율은 파종시기별(6월 20일, 7월 1일, 7월 10일, 7월 20일, 7월 30일, 8월 10일)로 93.3%~99.4% 범위였고, 중생종은 91.1%~96.7%, 중만생종 95.6%~100% 범위로 모든 생태형에서 적기파종(6월 20일)보다 늦게 파종할수록 출현율이 증가하는 경향을 나타내었다(Fig. 2).
출현 소요일수도 조생종, 중생종, 중만생종 모두 적기 파종시 5일이 소요되었으나 후기파종한 모든 처리에서 3일이 소요되어 출현 소요일수가 단축되었다. 개화 소요일수는 조생종의 경우 적기 파종시 개화 소요일수는 49일이었으며, 7월 1일 파종과 7월 10일 파종은 46일, 7월 20일 파종은 41일, 7월 30일 파종과 8월 10일 파종은 38일이 소요되어 적기파종 대비 개화 소요일수는 전체적으로 단축되는 경향을 보였다. 중생종과 중만생종도 조생종과 마찬가지로 적기파종 대비 늦게 파종할수록 개화 소요일수는 단축되는 경향을 나타내었다.

콩은 영양생장기와 생식생장기로 구분하는데, 이때 생식생장 단계는 일반적으로 R2를 기준으로 하고 있다(Lee et al., 2019; Chen and Wiatrak, 2010). 따라서 콩의 파종시기별 파종~개화기 단계(S~R2)와 개화기~수확기 단계(R2~R8)에 소요된 일수를 조사하였다. 파종 적기를 기점으로 일장이 짧아짐으로 파종기가 늦춰질수록 조생종, 중생종, 중만생종 모든 생태형에서 파종~개화기 소요일수가 단축되었고 개화기~수확기 단계도 적기보다 늦게 파종할수록 단축되는 경향을 나타내었다(Fig. 3). 그러나 중생종과 중만생종의 8월 10일 파종의 경우 꼬투리가 11월 6일(19.7℃)까지도 푸른색을 유지하고 있었고 11월 20일(7.8℃)에서는 다소 황색으로 등숙이 진행되었지만 11월 30일(3.1℃)에도 같은 색을 유지하고 있었고 콩도 더 이상 비대 되지 않아 성숙 후기 저온으로 인해 R5 단계에서 더 이상 성숙이 진행되지 않음을 확인할 수 있었다(Fig. 4).


파종시기별 온도와 일장 변화에 따른 생육반응
파종시기별 파종~개화기(R2), 개화기(R2)~수확기(R8)까지의 소요일수와 누적 일장 및 적산온도와의 관계를 조사하기 위해 다중회귀분석을 실시하였다(Tables 2, 3). 먼저 회귀모형의 유의성 검증을 위해 종속변수를 소요일수로, 적산온도와 누적일장을 독립변수로 회귀모형에 대한 ANOVA 분석을 실시한 결과 95% 유의수준에서 유의확률 값이 파종~개화기, 개화기~수확기까지 각각 P = 0.000, P = 0.000으로 0.05보다 적기 때문에 회귀식의 설명력이 유의한 것으로 나타났다(Table 1). 회귀분석에서 R2값은 독립변수인 온도와 일장이 종속변수인 소요일수에 얼마나 영향을 미치는지를 의미하는 것으로 파종에서 개화기까지의 R2값은 0.971, 개화부터 수확기까지는 0.979로 다중회귀모델이 두 기간 모두 97% 이상의 설명력을 가지는 것으로 나타났다. 온도와 일장이 개화 소요일수에 미치는 영향에 대해 다중회귀 분석을 실시한 결과, 유의확률은 일장이 0.013, 온도는 0.018로 P < 0.05 유의수준에서 유의한 것으로 나타나 일장과 온도 모두 개화 소요일수에 영향을 미치는 것으로 나타났다.
다중회귀 분석에서 비표준화 계수(B)는 독립변수(일장과 온도) 1이 증가할 때 종속변수(소요일수)가 얼마나 변화하는가를 의미하고 표준화 계수(β)는 변수들의 분포 정도를 계산한 것이므로 상대적으로 영향력을 비교할 수 있다. 파종에서 개화 소요일수까지의 표준화 계수는 일장은 0.511, 적산온도는 0.482로 개화 소요일수는 누적일장의 영향이 적산온도보다 더 큰 것으로 보인다(Table 2). 그리고 일장이 길수록 개화까지 소요일수는 길어지고, 적산온도도 높을수록 개화 소요일수가 길어지는 것으로 나타났다. 적기파종기인 6월 20일은 우리나라에서 일장이 최대가 되는 기간이고 이후 파종은 일장이 짧아지기 때문에 파종 적기보다 늦게 파종할수록 개화까지 소요일수가 단축되었고 이는 선행결과와 일치한다(Major et al., 1975). 따라서 적산온도와 누적일장이 개화 소요일수에 미치는 회귀모형은 개화 소요일수(Y) = 4.177 + (0.030 × 일장의 합) + (0.018 × 적산온도)로 나타났다(Table 2). 예를 들어 파종부터 개화기까지의 누적일장이 500 h이고, 적산온도가 1,000℃일 경우 개화까지 소요일수는 37일 정도 소요되는 것으로 나타났다. VIF값은 다중공선성을 나타내는 것으로 너무 비슷한 변수가 독립변수에 포함된 경우를 말하는 것으로 일반적으로 10 미만이면, 다중공선성에는 문제가 없다고 한다. 그러나 본 실험에서는 다중공선성이 17.203으로 매우 높아 다중공선성을 내포하고 있어 종속변수인 온도와 일장을 합쳐 수정된 회귀모형을 구하였다. 수정된 회귀모형은 개화 소요일수(Y) = 3.177 + (0.030 × (누적일장 + 적산온도))로 나타났다(Table 3).
Table 1.
ANOVA analysis of Multiple regression analysis model on the sum of temperature and day length on the number of growth days according to sowing times.
| Source of variation | dfy) | SSx) | MSw) | F value | P value |
|
No. of growing days (S~R2) MRz) model. | 2 | 306.561 | 153.281 | 254.055 | 0.000*** |
|
No. of growing days (R2~R8) MRz) model. | 2 | 1,483.086 | 741.543 | 341.984 | 0.000*** |
Table 2.
Multiple regression analysis of the sum of temperature and day length on the number of growing days according to sowing times.
| Dependent variable | Independent variable | Bz) | βy) | t value | P value | VIFx) | R2 |
|
No. of growing days (S~R2) | (Constant) | 4.177 | 0.971 | ||||
| Sum of day length | 0.030 | 0.511 | 2.816 | 0.013* | 17.203 | ||
| Sum of temp. | 0.018 | 0.482 | 2.659 | 0.018* | 17.203 | ||
|
No. of growing days (R2~R8) | (Constant) | 5.957 | 0.979 | ||||
| Sum of day length | 0.105 | 1.497 | 7.398 | 0.000*** | 28.627 | ||
| Sum of temp. | -0.017 | -0.522 | -2.579 | 0.021* | 28.627 |
Table 3.
Modified multiple regression model of the sum of temperature and day length on the number of growth days according to sowing times.
| Dependent variable | Independent variable | Bz) | βy) | t value | P value | VIFx) | R2 |
|
No. of growing days (S~R2) | (Constant) | 3.074 | 0.970 | ||||
|
Sum of day length + Sum of temp. | 0.023 | 0.001 | 22.908 | 0.000*** | 1.000 | ||
|
No. of growing days (R2~R8) | (Constant) | 20.945 | 0.964 | ||||
|
Sum of day length + Sum of temp. | 0.021 | 0.001 | 14.339 | 0.000*** | 1.000 |
개화기(R2)부터 수확기까지의 소요일수와 적산온도 및 누적 일장과의 관계를 다중회귀분석한 결과, 누적일장과 적산온도 모두 P < 0.05보다 적어 수확시기에 영향을 미치는 것으로 나타났다. R2값은 0.979로 97% 이상의 설명력을 나타내었고, β값은 누적 일장이 1.497, 적산온도가 –0.522로 일장은 정의 상관, 온도는 부의 상관을 나타내었다. Nico et al. (2015)은 개화기부터 수확기까지의 일장이 짧아지면 등숙기간이 짧아져 수량이 감소한다고 하였고, Thuzar (2010)은 등숙기의 온도가 높아질수록 등숙이 빨리 진행되어 성숙 및 수확시기가 단축된다고 하여 일장은 생육일수와 정의 상관, 온도는 부의 상관이 있어 수확시기가 단축되는 것으로 보인다. 따라서 온도와 일장이 개화기부터 수확기까지 소요일수에 미치는 회귀모형은 개화기~수확기까지 소요일수 (Y) = 5.957 + (0.105 × 누적일장) ‒ (0.017 × 적산온도)로 나타났고 다중공선성으로 인한 수정된 회귀모형은 수확일수(Y) = 20.945 + (0.021 × (누적일장 + 적산온도))으로 나타났다. 수정 전 회귀모형과 수정 후 회귀 모형을 앞선 예시(누적일장 500 h과 적산온도 1,000℃)로 비교해 보면 파종~개화기, 개화기~수확기까지의 소요일수는 수정된 회귀모형에서 각각 11일, 12일이 길어지는 경향을 나타내었다. 그러나 수정전 회귀모형에서 다중 회귀분석의 목적이 개별변수에 대한 추정이 아닌 종속변수(소요일수) 전체에 대한 예측일 경우 다중공선성은 문제가 되지 않는다는 견해도 있다(Grewal et al., 2004; Alin, 2010). 따라서 위의 수정전․후 회귀모형은 실제 생육일수와의 비교를 통해 검정 및 수정이 필요할 것으로 보인다. 즉 고온에 의한 개화 지연, 성숙기 저온와 같은 생태적 특성을 비롯하여 콩 생태형에 따른 지역별, 시기별 파종시기 및 기상 자료를 활용한 다양한 변수에 대한 분석이 필요할 것으로 보인다.
생육, 수량구성요소 및 수량
파종시기별 생육과 수량구성요소 변화를 조사한 결과 모든 생태형에서 경장, 마디수, 분지수는 적기파종에서 가장 길었고 적기보다 늦게 파종할수록 줄어드는 경향을 나타내었다(Table 4). Park et al. (2014)은 조생종을 만파 할수록 마디수가 증가한다고 하여 본 연구와 상반되었고, 중․만생종의 경우 만파 할수록 마디수가 감소한다는 보고와는 일치하였다. 수량구성 요소인 협수도 모든 생태형에서 적기파종 처리에서 가장 높았고 적기 파종 대비 늦게 파종할수록 감소하였다. 협당립수는 모든 생태형에서 8월 10일 처리구에서 가장 낮았고, 적기 파종보다 늦게 파종할수록 다소 낮아지는 경향이었으나 통계적으로 유의한 차이는 없었다.
Table 4.
Comparisons of growth parameters and yield components according to six sowing times with soybean ecotype.
| Ecotype |
Sowing date (Month. day) |
Stem length (cm) |
Stem diameter (mm) |
No. of node (ea/plant) |
No. of branch (ea/plant) |
No. of pod (ea/plant) |
No. of seed (ea/pod) |
|
Early maturing | 6.20 | 72.5az) | 9.5a | 15.5a | 4.5a | 69.3a | 2.2a |
| 7.1 | 71.9a | 9.2ab | 14.4ab | 4.4a | 67.6a | 2.0ab | |
| 7.10 | 67.8ab | 8.9ab | 14.3ab | 3.3ab | 63.6a | 1.8b | |
| 7.20 | 67.7ab | 8.2ac | 14.0b | 3.3ab | 55.4ab | 1.8ab | |
| 7.30 | 65.1b | 7.9bc | 14.1b | 2.8b | 41.3b | 1.8b | |
| 8.10 | 57.9c | 7.2c | 12.5c | 2.8b | 11.5c | 0.7c | |
|
Middle maturing | 6.20 | 81.1a | 10.0a | 15.4a | 5.9a | 77.0a | 2.0a |
| 7.1 | 73.2b | 8.7b | 15.3a | 5.1a | 78.6a | 2.0ab | |
| 7.10 | 72.8b | 8.3b | 15.1a | 3.7b | 74.4a | 1.8ac | |
| 7.20 | 72.2b | 8.4b | 14.1b | 3.2bc | 54.5b | 1.7bc | |
| 7.30 | 71.9b | 8.1b | 14.0b | 2.4c | 49.4b | 1.7c | |
| 8.10 | 57.0c | 6.3c | 12.1c | 3.2bc | 18.3c | 1.4d | |
|
Mid-late maturing | 6.20 | 62.6a | 9.7a | 13.2a | 6.0a | 77.3a | 2.0a |
| 7.1 | 59.9ab | 9.3a | 11.6b | 5.7a | 64.1b | 1.9a | |
| 7.10 | 59.1ac | 9.3a | 11.1bc | 3.9b | 58.3b | 2.0a | |
| 7.20 | 55.5bc | 7.4b | 10.5c | 3.1bc | 42.7c | 2.0a | |
| 7.30 | 52.7cd | 6.9b | 10.5c | 2.8c | 38.9c | 2.0a | |
| 8.10 | 47.3d | 5.8c | 10.5c | 3.3bc | 24.7d | 1.7b | |
| Ecotype (E) | 0.000*** | 0.135ns | 0.000*** | 0.011* | 0.016* | 0.000*** | |
| Sowing date (S) | 0.000*** | 0.000*** | 0.000*** | 0.000*** | 0.000*** | 0.000*** | |
| E×S | 0.193ns | 0.049* | 0.018* | 0.221ns | 0.261ns | 0.000*** | |
백립중은 조생종의 경우 적기파종(6월 20일)에서 7월 20일 파종까지는 큰 차이가 없었으나 7월 30일, 8월 10일 파종은 급격히 감소하였다(Fig. 5). 중생종과 중만생종은 7월 10일 파종까지는 큰 차이가 없었으나 7월 20일 파종부터 감소하는 경향을 나타내었고, 8월 10일 파종은 매우 낮은 백립중을 나타내었다. 수량도 모든 생태형에서 적기파종에서 가장 높았고 이후 파종시기가 늦어질수록 수량이 감소하였다. 모든 생태형에서 7월 20일 이후 파종에서 수량이 급격히 감소하는 경향을 나타내었는데, 후기파종시 적기 파종대비 조생종은 27%, 중생종은 49%, 중만생종은 38%의 수량 감소를 보였다. 그리고 7월 30일 파종과 8월 10일 파종은 수량이 급격히 감소하였는데 이것은 저온으로 인해 등숙이 더 이상 진행되지 않아 수량이 매우 낮았기 때문이다(Fig. 4).
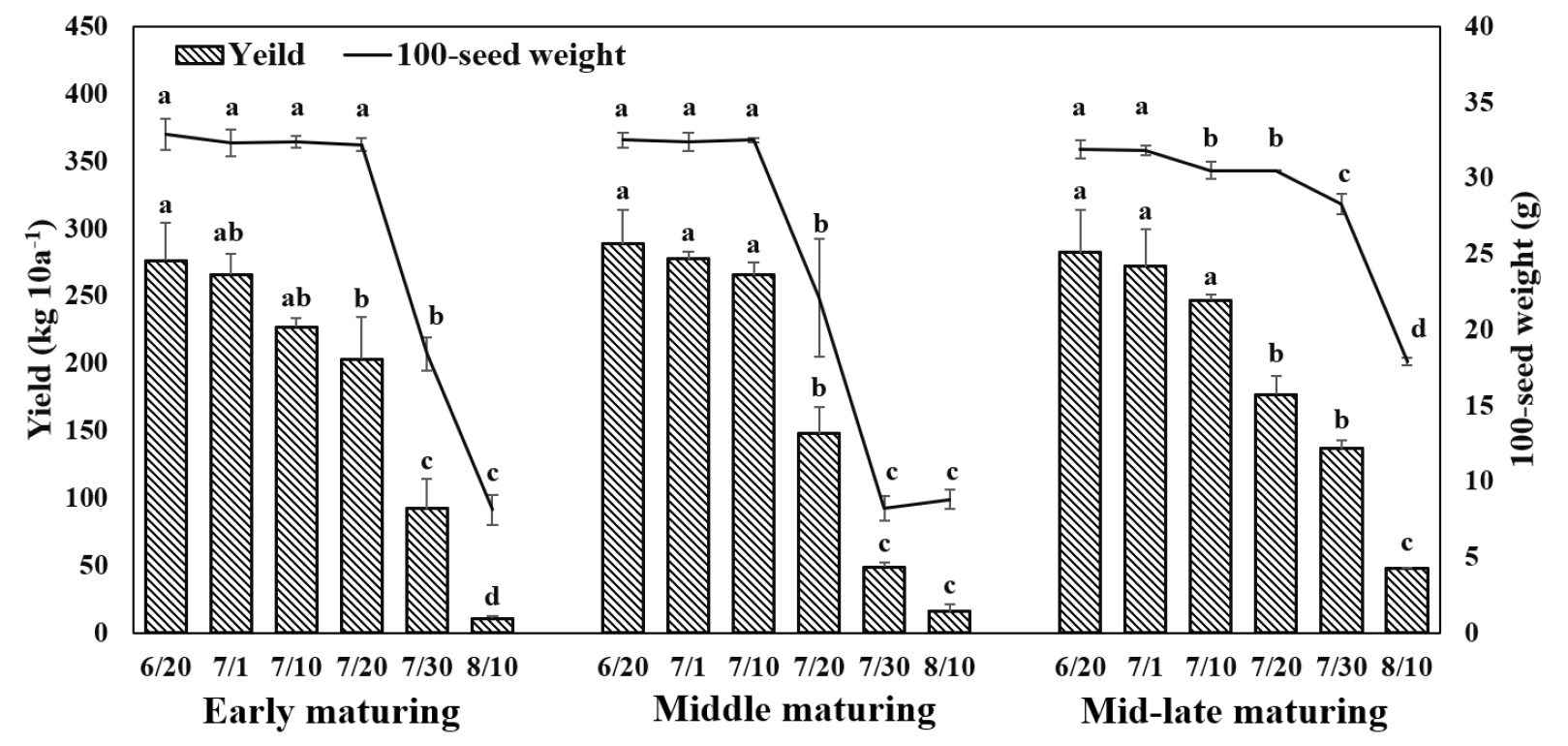
적산온도와 누적일장이 수량에 미치는 영향을 조사하고자 다중회귀분석을 실시하였으나 유의한 차이를 볼 수 없었다. 이것은 수량이 온도와 일장 간 상관성이 크고 두 요인 외에 다양한 환경적인 요인(토양비옥도, 수분함량 등)이 관여하기 때문인 것으로 보인다(Córdova et al., 2020; Wei et al., 2020; Mandić et al., 2017). 파종시기에 따른 생육일수는 일장과 온도와 각각 R2 = 0.884, R2 = 0.8759로 밀접한 상관이 있기 때문에 생육일수와 단위수량 및 수량구성요소 간 상관관계를 분석하였다(Figs. 6, 7). 생육일수는 협수(r = 0.9), 협당립수(r = 0.7), 백립중(r = 0.8), 수량(r = 0.9), 경태(r = 0.8), 분지수(r = 0.7)와는 밀접한 관련이 있었고, 초장은 중 정도의 상관(r = 0.5), 마디수(r = 0.3)는 낮은 상관을 나타내었다. 초장과 마디수는 콩의 생육일수와 밀접한 관련이 있지만 본 실험의 공시 품종 중 조생종인 새올 품종은 일장이나 온도에 둔한 품종으로 3품종을 모두 합쳐서 분석하였기 때문에 나타난 결과로 보인다(Kim et al., 2023). 그리고 수량은 생육일수(r = 0.9)나 협수(r = 0.9), 협당립수(r = 0.7), 백립중(r = 0.9) 등 수량구성요소와는 밀접한 관련이 있었고, 생육지표 중 경태(r = 0.8)과 분지수(r = 0.7)와도 높은 상관을 나타낸 반면, 경장(r = 0.5)과 마디수(r = 0.4)와는 중정도의 상관을 나타내었다. 따라서 파종시기에 따라 생육일수가 달라지는데 후기파종으로 적정 생육일수가 감소할수록 수량을 포함한 수량구성요소들이 감소하는 것으로 나타났기 때문에 적기 파종 이후에는 생육 및 수량이 감소함을 확인할 수 있었다.
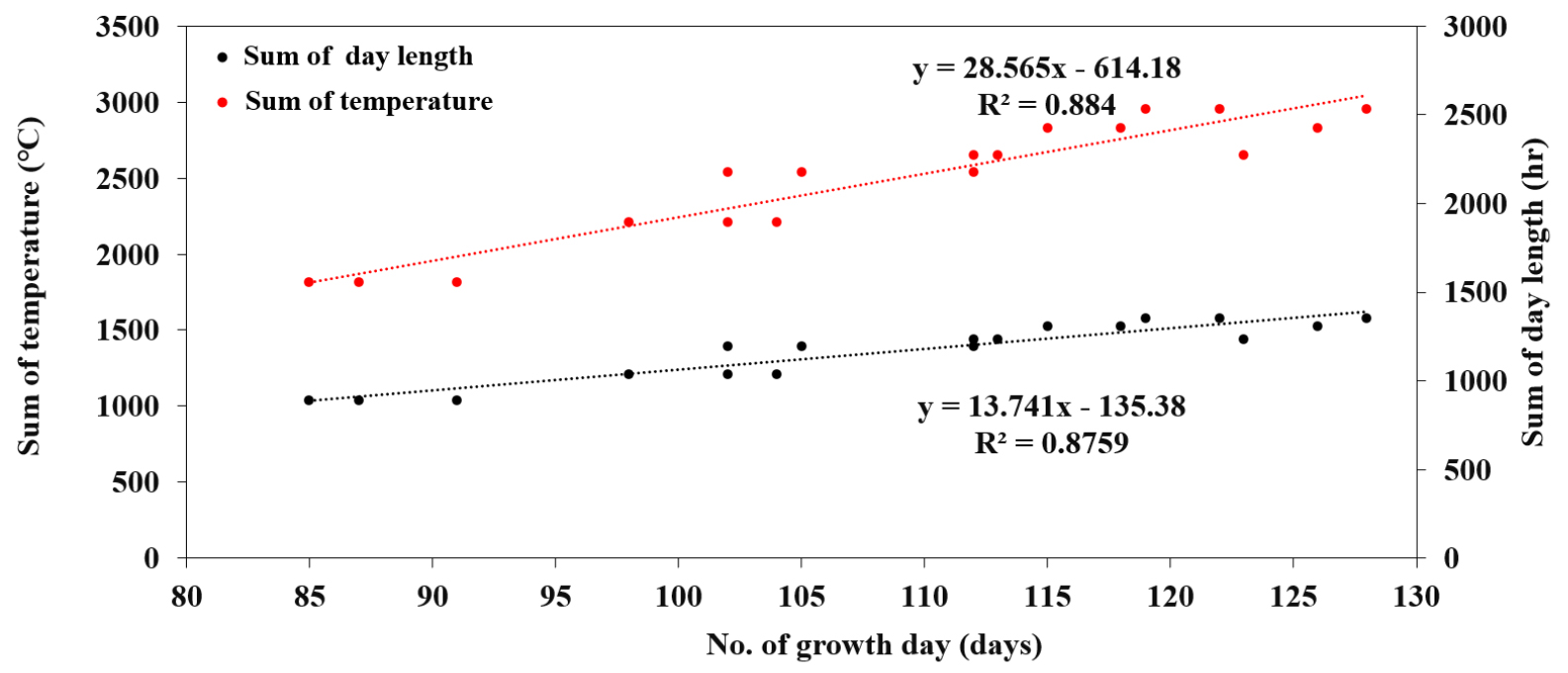

Fig. 7.
Pearson’s correlation coefficients of GD (growth day) with yield component elements. SL (steam length), SD (stem diameter), NB (number of branch), NM (number of main node), NP (number of pods), NSP (number of seeds per pod), HSW (hundred seed weight), Y (yield). *,**,***denotes statistically significant values p < 0.05, 0.01, and 0.001, respectively.
적 요
남부지방(진주)에서 콩 생태형에 따라 적기파종 이후 파종 시기를 달리하였을 때 온도와 일장이 생육일수 및 수량에 미치는 영향을 조사한 결과는 다음과 같다.
1. 적기파종 이후 파종시기를 달리하여 파종하였을 때 모든 생태형에서 출현일, 개화소요일수, 전체 생육일수는 단축되었다.
2. 일장과 온도가 생육일수에 미치는 영향을 다중회귀분석으로 분석한 결과 파종~개화기까지 소요일수는 일장과 온도 모두 정의 상관이였고, 개화기~수확기까지 소요일수는 일장은 정의 상관, 온도는 부의 상관이 있었다.
3. 추정 회귀모형은 파종~개화까지 소요일수(Y) = 3.177 + (0.030 × (누적일장 + 적산온도)), 개화기~수확기까지 소요일수는 (Y) = 20.945 + (0.021 × (누적일장+적산온도))인 것으로 나타났다.
4. 수량은 적기파종에서 가장 높았고 적기파종 이후 늦게 파종할수록 수량이 급격히 감소하여 조생종은 7월 20일, 중만생종은 7월 10일 이전까지는 파종을 해야 급격한 수량감소를 피할 수 있을 것으로 보인다.
