

Maize (Zea mays L.) is one of the most important cereals in the world, but global climate change adverse effects maize production in various ways (Erdal et al., 2015). Among abiotic stresses, drought has significant influence on the various stage of maize life cycle such as seedling, vegetative and reproductive growth. It has been shown baneful impacts on leaf expansion, stem elongation, root proliferation, water use efficiency, metabolism, enzyme activity, ionic balance, solute accumulation, and chlorophyll contents (Khan et al., 1999; Farooq et al., 2002). Ultimately, drought stress (DS) causes reduction in grain yield (Athar and Ashraf, 2005; Beck et al., 2007; Araus et al., 2008; Farooq et al., 2009). However, responding toward drought stress, various adaptive strategies have been employed in morphology, biochemistry, and physiology (Lu et al., 2011).
Maize grain yield is determined by many factors such as plant height (PH), leaf senescence (LS), tassel branch (TB), anthesis-silking interval (ASI), and ears per plant (EPP) (Fuad-Hassan et al., 2008). When water shortage happens before flowering, it does have extreme harmful influence on anthesis, thus silk emergence from husks is delayed, and causes an increased ASI (Bassetti and Westgate, 1993b; Edmeades et al., 2000). In many studies, the sensitivity of silk growth toward water shortage has been defined as the growth rate of silk emergence (Herrero and Johnson, 1981; Westgate and Boyer, 1985; Bassetti and Westgate, 1993b). Increased ASI ultimately leads to low grain yield (Edmeades et al., 1992). For this reason, ASI plays important roles as drought tolerance index (TI). Drought tolerance has been adapted for diminishing ASI in maize (Edmeades et al., 1993; Bolaños and Edmeades, 1996; Bruce et al., 2002). A short ASI is correlated with quantitative trait loci (QTLs) of increased grain yield (Ribaut et al., 1997). The maintenance of rapid silk growth is genetically linked to the conservation of grain yield under drought stress.
Tropical maize has been studied over the 40 years to enhance drought tolerance in breeding programs. The international Maize and Wheat Improvement Center (CIMMYT) had developed drought tolerant genotypes through recurrent selection and evaluation under managed stress conditions in the 1970s (Fischer et al., 1989; Bolaños and Edmeades, 1993; Byrne et al., 1995; Edmeades et al., 1999). CIMMYT developed several inbred lines from these drought tolerant lines, and released hybrids that possess high grain yield potential in the tropical climate (Beck et al., 1996). Regarding temperate maize, genetic modification methods have been developed in parallel with traditional breeding to improve drought tolerance, particularly in the US Corn Belt (Cooper et al., 2014). The objectives of this study were to evaluate for drought tolerance and identify the drought tolerant genotypes.
MATERIALS & METHODS
Plant materials
Maize lines were selected from different sources, nine inbred genotypes from USDA (B73, CML103, CML228, CML277, CML322, CML69, Ki3, Ki11, and NC350), three Southeast Asian genotypes (DK9955, LVN-4, and 333), and five Korean hybrids (Cheongdaok, Gangdaok, Ilmichal, Kwangpyeongok, and Pyeonganok). Drought stress responses among experimental maize lines were determined in the Dongguk university greenhouse facility.
Plant growth conditions
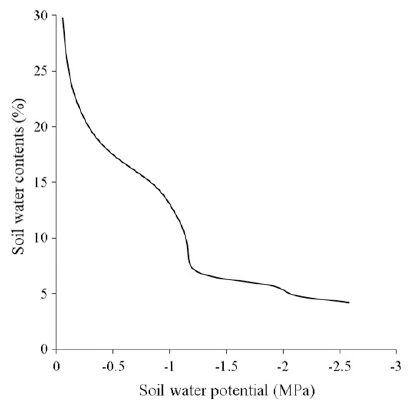
Three seeds were planted onto a plastic pot 37 cm wide and 37.5 cm high (20 L) filled with soil (Plant world, Nongwoo bio) on April. Repeated experiments were done in the year 2015 and 2016. Plants were thinned to one plant per pot at the V3 stage. At the time prior to the tassel emergence, water supply was withheld until soil water content was around 7% (v/v) at day 4~5 post water-withholding. We measured soil water content (%) and soil water potential (MPa). Fig. 1 shows the correlation between soil water content and soil water potential. Water was re-irrigated to the plants at day 20 post water-withholding. The well-watered (WW) pots were kept over 20% (v/v) of soil water content. Volumetric soil water content was measured with a FDR2 (Frequency Domain Reflectometry) type sensor (WT-1000N, Mirae sensor, Korea). Soil water potential was measured with dielectric water potential sensors (MPS-6, Decagon Devices, Pullman, WA, USA). At each pot, one sensor located at a depth of 15 cm and connected to the EM50 data loggers (Decagon Devices). Fertilizer was applied at 20 : 15 : 15 kg/10 a of N : P2O5 : K2O, respectively. Additionally, 10 kg/10 a of nitrogen was applied at day 45 post planting.
Data collection
We scored date of anthesis and silking when pollen shedding and extruded silk are visible, then calculated as the day to silking minus day to anthesis for each plant (Buckler et al., 2009). Leaf senescence (LS) was scored using a scale from 0 to 10 at silking date, estimated total leaf area is dead by 10 (1 = 10% dead leaf area; 2 = 20% dead leaf area; 3 = 30% dead leaf area; 4 = 40% dead leaf area; 5 = 50% dead leaf area; 6 = 60% dead leaf area; 7 = 70% dead leaf area; 8 = 80% dead leaf area; 9 = 90% dead leaf area; 10 = 100% dead leaf area). To determine ears per plant (EPP), the number of ears was divided by the number of each plant (Bänzinger et al., 2000). Tassel length (TL) was measured as the length between base of tassel and the top of tassel at silking date. Fresh weight (FW) was measured as total aerial part at silking date.
RESULTS & DISCUSSION
Traits for drought tolerance
Table 1 represents the results of the analysis of variance (ANOVA) for ASI, LS, EPP, TL, and FW under WW and DS conditions. ANOVA analysis revealed highly significant difference (p <0.01) among genotypes in all tested traits. Mean value and standard deviation (SD) confirms phenotypic variation among genotypes. We analyzed drought index following CIMMYT-derived drought tolerance (Ki11) and susceptibility (Ki3) (Prasanna et al., 2010).
Table 1
Analysis of variation in ASI, leaf senescence, ear per plant, tassel length and fresh weight of different maize genotypes under WW and DS conditions at 2015.
Values followed by the identical letters in the row are not significantly different.
Values are means ± SE; ASI = anthesis-silking interval; DS = drought-stress; WW = well-watered.
Anther-silking interval (ASI)
Under DS condition, the mean values for ASI are significantly increased that compared to the WW condition. ASI of Ki11 and Ki3 is 0.3/2.3 and 2.9/12.9 days under the WW and DS conditions, respectively. Ilmichal, CML277, LVN-4, DK9955, CML69, 333, Ki3, Pyeonganok, and Gangdaok are recorded to more than 6 days under DS conditions. ASI is a major secondary trait for the drought tolerance selection in maize (Bolaños and Edmeades 1993; Edmeades et al., 1993; Byrne et al., 1995; Bolaños and Edmeades 1996; Ribaut et al., 1997; Bänzinger et al., 2000; Ziyomo and Bernardo, 2012). Previous study reported silk senescence occurred following a few days after first silk appearance, and the flower function loss happened 4 to 10 days after the first silks emergence according to genotypes (Bassetti and Westgate 1993a). Ilmichal, CML277, LVN-4, DK9955, CML69, 333, Ki3, Pyeonganok, and Gangdaok showed the decreased pollination rate under DS condition. Rowland (1993) reported a few days of retardation of anthesis in days among genotypes under drought stress. Angus and Moncur (1997) and Morgan (1980) presented similar report. According to Rowland (1993), drought stress at silking stage interrupted silk emergences from the cob husk, caused silks dryness, and inhibited pollen tube growth. Drought stress at vegetative, flowering, and reproductive stage led to reduction in grain yield (Bawa et al., 2015).
Leaf senescence
We observed leaf senescence under the WW and DS conditions. Leaf senescence was elevated in all genotypes (except CML228). This result is coinciding with the report of Quarrie and Jone (1977). Quarrie and Jone (1977) found that drought stress impacts on leaf development and vegetation. Inhibition of leaf development resulted in the reduction of cell expansion and cell division under drought stress. In addition, drought stress caused reduction in the leaf area, radiation use efficiency, and harvest index by overproducing ROS and accelerated leaf senescence (Nogués and Baker, 2000; Earl and Davis, 2003). In this study, difference between WW and DS in Ilmichal, CML103, and Ki11 is less than 1, otherwise more than 1.5 in Pyeonganok, Cheongdaok, Ki3, CML322, CML69, DK9955, and 333. According to Lawlor and Cornic (2002), low relative water contents decreased leaf water potential and photosynthetic metabolism. Leaf senescence was inversely correlated to grain yield. Grain filling needs nitrogen uptake, however higher leaf senescence limits nitrogen supply (Masclaux-Daubresse et al., 2010).
Ears per plant (EPP)
Regarding EPP, Ki11, CML277, CML69, Pyeonganok, CML103, and Cheondaok have over 0.5 decreases, on the other hand, Gangdaok shows more than 0.5 increases. EPP is related to grain yield. Grain yield is ultimate goal for drought tolerance. EPP is an important factor conferring drought tolerance, but EPP was not linked to drought tendency in reference.
Tassel length (TL)
Tassel length has an influence on grain yield in drought condition. Hunter (1969) reported the negative effect of the tassel on grain yield with 19 percent reduction. In this study, we found more than 3 cm increase in NC350, CML277, and CML228, in contrast to more than 1 cm decrease in Ki11, Kwangpyeongok, Ki3, Gangdaok, CML69, and LVN-4.
Fresh weight (FW)
Regarding FW, Ki11, CML69, Ilmichal, LVN-4, DK9955, and Pyeonganok showed over 150 g declines, however Ki3, NC350, and 333 represented under 100 g reduction in shoot weight. This result is not corresponds to the reference. In Ashraf’s (1989) report, drought tolerant lines had higher shoot weight than susceptible genotypes.
ASI analysis for drought tolerance
ASI and LS are similar to drought tendency of Ki11 and Ki3, however EPP, TL, and FW are different. According to ASI and LS, CML103 and Ki11 are drought tolerant genotypes, whereas Ki3 and 333 are drought susceptible. Grain yield is correlated strongly to EPP, ASI, but moderately to LS. (Smith et al., 2004). Therefore, we repeated ASI experiment (Fig. 2). ASI of Ki11 and Ki3 is 2.5/4.0 and 2.1/8.6 days under WW and DS conditions in 2016 experiment, respectively (Fig. 2a, c). ASI of CML228, CML103, Cheongdaok, NC350, B73, Ki11, CML322, and Kwangpyeongok are recorded less than 4 days under WW and DS conditions in 2016 (Fig. 2a). On the other hand, ASI of CML277, LVN-4, DK9955, CML69, 333, Ki3, Pyeonganok, and Gangdaok are more than 4 days under DS conditions (Fig. 2b, c). According to Bolaños and Edmeades’s (1996) report, ASI of six tropical maize species was 2.3 and 8.3 days under well-watered and severe water stress conditions, respectively. ASI of CML103, Ki11, CML322, Kwangpyeongok, CML277, LVN-4, Ki3, Pyeonganok, and Gangdaok were similar, whereas ASI of CML228, Cheongdaok, NC350, B73, Ilmichal, DK9955, CML69, and 333 were different in 2015 and 2016. These results implicate influence of environmental factor.
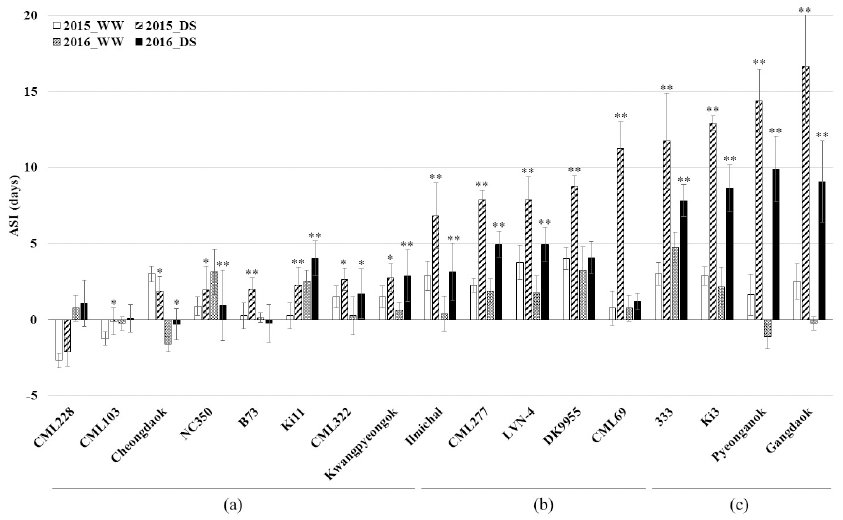
Fig. 2
Mean and standard deviation of ASI (days) for maize genotypes under well-watered (WW) and drought-stress (DS) conditions. (a) Low range of ASI (< 5 days), (b) medium range of ASI, and (c) high range of ASI (> 7 days). (Duncan’s tests, *p < 0.05, **p < 0.01).ASI of Ki3, Pyeonganok, and Gangdaok is different over 5 days between DS and WW conditions in 2015 and 2016 (Fig. 3). On the other hand, ASI of CML228, CML103, Cheongdaok, NC350, B73, Ki11, CML322, and Kwangpyeongok is different less than 3 days between DS and WW conditions in 2015 and 2016. According to Udomprasert et al’s (2005) report, drought stress reduced photosynthesis, silk growth, but increased ASI during tasseling time. As a result, drought stress causes a reduction in corn kernel set. Udomprasert et al. (2005) evaluated greater tolerance in Ki11 than Ki3 under DS conditions. According to Andersen et al. (2002), drought stress caused an increased ASI about 3 to 7 days. In addition, drought stress impacted on ASI with increased about 4.5 days compared to average of 1 day under WW conditions (Chapman and Edmeades, 1999). Hall et al. (1981) reported that ASI increase from 3 to 11 days was inversely related to grain yield of 71% reduction. Moss and Downey (1971) reported similar results. Drought tolerant maize was selected by referencing on ASI reduction (Bolaños and Edmeades, 1993). Grain yield reduction was correlated to impediment of kernel development and filling (Shin et al., 2015). Kernel development and filling were inhibited under severe water stress at flowering stage (Robins and Domingo, 1953; Moss and Downey, 1971; Schussler and Westgate, 1991a and b; Setter et al., 2001). Eight days of drought stress at flowering stage caused reduction of the 100-grain weight about 20% (Grant et al., 1989). The 100-grain weight was significantly reduced under severe water stress condition after silk emergence, however not significantly different under mild water stress (Harder et al., 1982). In this study, the maize genotype was evaluated as ASI of less than 5 days under DS condition and less than 3 days between DS and WW conditions were classified to drought tolerant. On the other hand, the drought susceptible genotypes determined as ASI of more than 7 days under DS condition and more than 5 days between DS and WW conditions.
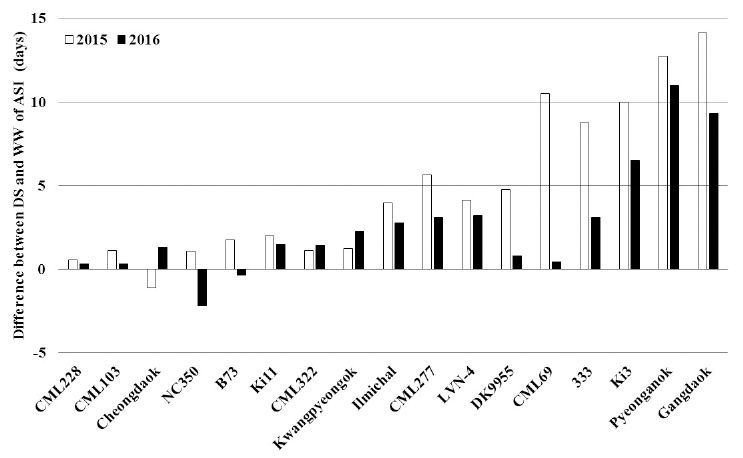
Fig. 3
Difference in ASI between drought-stress (DS) and well-watered (WW) conditions of the maize genotypes.ASI analysis showed that CML228, CML103, Cheongdaok, NC350, B73, Ki11, CML322, and Kwangpyeongok are drought tolerant genotypes, whereas Ki3, Pyeonganok, and Gangdaok are drought susceptible. Although B73 is reported as drought susceptible genotype, ASI of B73 determined less than 5 days under DS condition and less than 1.5 days between DS and WW conditions (Sari-Gorla et al., 1999; Gonzalo et al., 2006). In other words, ASI of B73 is not significantly affected by drought stress. We suggest that ASI for drought tolerance screening is a practical method. To further confirm the relationship between ASI and grain yield, extended studies are required.
CONCLUSION
We examined ASI, LS, EPP, TL, and FW under WW and DS conditions. ASI and LS results are compatible with drought tendency in references. Other traits (EPP, TL, and FW) are different from references. According to ASI and LS analysis, CML103 and Ki11 are drought tolerant genotypes, otherwise Ki3 and 333 are susceptible genotypes. Grain yield is correlated strongly to EPP, ASI, but moderately to LS. ASI of most genotypes is similar in 2015 and 2016. Grain yield reduction coincides with high ASI score. Difference in ASI between DS and WW conditions of Ki3, Pyeonganok, and Gangdaok is over 6.5 days. On the other hand, ASI of CML228, CML103, Cheongdaok, NC350, B73, Ki11, CML322, and Kwangpyeongok is less than 3 days. With these results, we suggest that CML228, CML103, Cheongdaok, NC350, Ki11, CML322, and Kwangpyeongok are drought tolerant genotypes, otherwise Ki3, Pyeonganok, and Gangdaok are susceptible genotypes. These results contain the basic information regarding drought tolerance in maize.
