

서 론
재료 및 방법
시힘품종
몰리브덴 엽면 시비 처리
몰리브덴 고형비료 시비 방법
수발아 검정방법
수량 및 수량구성요소 조사
Total RNA 추출 및 Quantitation Real-Time PCR 분석
통계처리
결과 및 고찰
몰리브덴 처리 시기에 따른 수발아 경감 효과
몰리브덴 비료 처리량에 따른 수발아 영향
비료처리일에서 출수기까지 경과일수와 몰리브덴 농도별 수발아 차이
몰리브덴 처리에 따른 ABA 대사 관련 유전자 발현 분석
몰리브덴 처리에 따른 수량구성요소 변화
적 요
서 론
수발아(Pre-Harvest Sprouting, PHS)는 수확 전 고온 다습한 환경에서 이삭의 낟알에 싹이 트는 현상을 말한다(Park & Park, 1984). 수발아가 발생한 벼는 쌀의 품질 및 생산성을 떨어뜨릴 뿐만 아니라(Kim et al., 2008) 종자의 품질에도 크게 영향을 미친다. 수발아 피해립율이 높은 종자는 저장 시 온도와 습도가 높을수록 건전 종자에 비해 종자활력이 저하되고 미발아율과 부패율이 높아져 출아율에도 영향을 미친다(Shon et al., 2014). 수발아는 종자의 휴면성과 관계가 큰데, 벼는 주로 Abscisic Acid (ABA)/Gibberellic Acid (GA) 비율의 균형을 통해 종자 휴면이 조절된다고 알려졌다(Finch-Savage & Leubner-Metzger, 2006; Liao et al., 2018). ABA는 종자 성숙과 휴면에 관련된 물질로 알려져 있으며(Bewley & Black, 1994) ABA함량과 품종별 수발아성 사이에 부의 상관관계가 있다고 하였다(Liu et al., 2014).
수발아 피해 경감을 위해 품종 개발과 재배적 측면에서 다양한 방법들이 시도되어 왔다. 품종 개발 측면에서는 벼 종자의 휴면과 관련된 QTL이 벼의 재배종과 야생종 및 잡초성벼(weedy rice)의 염색체에서 확인되었고 QTL중 일부가 수발아성과 밀접한 관련이 있는 것으로 나타났다(Cai & Morishima, 2002; Gu et al., 2004; Lu & Snow, 2005). 또한 수발아 저항성 유전자인 OsPHS1 (Oryza sativa pre-harvest sprouting 1)가 보고되었다(Youn et al., 2016). 우리나라에서는 조생종이 중만생종보다 수발아가 발생하기 쉽다고 알려져 왔으나(Park & Park, 1984) 이는 품종적 특성보다는 출수기가 빠를수록 고온시기에 노출되어 등숙이 빨라지는 생태적인 특성으로 보고되었다(Kang et al., 2018). 따라서 재배적 측면에서 수발아를 회피하기 위해 출수기를 늦추어 낮은 온도에서 등숙이 이루어지도록 재배 시기를 조절하는 것이 효과적이다(Hwang et al., 2021).
그러나 벼에서 수발아 발생을 억제하는 처리제는 없는데 기후온난화로 인해 등숙기 기온이 상승함에 따라 수발아에 적극 대응할 수 있는 방법이 필요하다. 특히 자포니카벼는 인디카벼 보다 수발아가 발생하기 쉽고 최근 보급되고 있는 가루쌀은 수발아율이 높아 대책이 필요한 실정이다. 미량원소인 몰리브덴은 밀과 옥수수 등에서 수발아 감소 효과를 보인다는 보고가 있다. 옥수수는 몰리브덴 분무처리시 종자의 몰리브덴 농도가 증가하고 수발아가 감소했으며(Tanner 1978), 밀에서는 500 ppm의 몰리브덴 처리시 종자 발아율이 감소하고 ABA 함량은 증가하였다고 한다(Cairns et al., 1997). 벼의 몰리브덴 효과는 비교적 최근에 보고되었는데 지엽출현기에 몰리브덴 처리 시 수발아율이 감소 되었다고 한다(Tejakhod & Hammond, 2018).
몰리브덴은 ABA 생합성과정 중 보조인자(MOCOS, MCSU)로 관여하는 원소로 알려졌다(Mendel, 2013; Zhang et al., 2016). 애기장대의 ABA생합성 과정에서 최종산물을 활성화하는 AAO (Arabidopsis aldehyde oxidase) 유전자는 MoCo (Molybdenum Cofactor)에 의해 활성화된다(Watanabe et al., 2018). 벼에서 OsAO3 (Oryza sativa Aldehyde Oxidase 3) 과발현 돌연변이체는 종자 발아가 지연되고 유아 성장이 감소하였는데 OsAO3가 종자 발아 및 유아 생장을 조절에 관여하는 것으로 보고되었다(Shi et al., 2021). 하지만 벼에서 몰리브덴이 수발아에 미치는 영향에 대한 보고는 매우 적은편이다. 따라서 본 연구는 수발아 경감 효과를 나타내는 몰리브덴의 최적 처리 시기와 처리량을 구명하고, ABA와 관련된 유전자 발현 분석을 통해 몰리브덴의 생리기작을 밝히고자 하였다.
재료 및 방법
시힘품종
가루쌀 품종인 바로미2와 일반품종인 신동진을 사용하였다.
몰리브덴 엽면 시비 처리
몰리브덴 시비는 몰리브덴산염 수용액을 엽면시비 처리한 방법과 남해화학에서 제작한 NPK비료와 혼합한 입상형태 두 가지 방법으로 처리하였다. 먼저, 엽면시비처리는 국립식량과학원 인공기상동 유리온실에서 2023년에 수행하였다. 벼는 포트당 1주 1본씩 3본을 이앙하여 출수 전까지 24±5℃로 처리하였고 출수 이후 수확기까지 21±5℃로 처리하였다. 엽면시비는 증류수에 몰리브덴산염을 500, 1000 ppm으로 용해시켜 출수 전 15일과 출수 후 20일 두 시기에 포트당 3 ml씩 잎에 분무하였다.
몰리브덴 고형비료 시비 방법
고형비료 시험은 국립식량과학원 인공기상동 유리온실과 국립식량과학원 논포장에서 2024년에 각각 수행하였다. 질소-인산-가리 시비는 N-P205-K20 성분기준으로 9-4.5-5.7 (kg/10a)을 시용하였다. 몰리브덴 고형비료는 N-P205-K20 (18-1-15%) 입상비료에 몰리브덴산염(MoO42-)을 0.035, 0.07, 0.17%첨가하여 남해화학 농자재연구팀에서 제조하였다. 유수가 8 cm되는 시점을 출수 전 15일로 보고 몰리브덴 입상비료를 포트당 3.6 g씩 처리했으며 몰리브덴산염(MoO42-)함량으로 10a당 0, 21, 42, 102 g에 해당한다. 포장실험에서는 몰리브덴 입상비료를 N-P205-K20 (18-1-15%)에 몰리브덴산염(MoO42-)을 0.1, 0.2, 0.4% 첨가하여 남해화학에서 제조하였다. 제조한 NPK-Mo 비료를 유수형성 초기인 출수 전 25일에 각각 11kg/10a로 처리하였는데, 이는 MoO42-함량 기준으로 10a당 0, 11, 21, 42 g에 해당한다.
수발아 검정방법
수발아 검정방법은 출수 후 적산온도 1,000도에서 채취한 이삭을 25℃ 항온에서 4일간 처리 후 수발아조사한 방법과, 22±5℃ 변온조건에서 4일간 분무처리한 두 가지 방법으로 조사하였다. 등숙이 동일한 시점에 수발아를 평가하기 위해 이삭별로 출수일을 표시하였다. 인공기상동 포트 시험은 출수 후 적산온도 1,000도에서 이삭을 채취하였는데, 엽면시비는 변온조건에서 분무처리 방법으로 수발아 검정을 하였고 고형비료 시비는 분무처리와 항온항습 방법으로 검정하였다. 포장 시험은 출수 후 적산온도 1,200도에서 수확한 후 제현하여 시험구당 현미 15 g씩 3반복에 대한 발아립율을 조사하였다. 수발아율(%)은 (발아립수/이삭의 총립수) ×100으로 계산하였다.
수량 및 수량구성요소 조사
포장시험은 처리별 4.5 m2 면적으로 3반복구로 수확하여 정조 및 현미의 수량을 조사하였다. 포장시험 및 포트시험에서 수량구성요소는 출수 후 적산온도가 1,200℃되는 시기에 3주씩 3반복으로 이삭당 영화수, 등숙률, 천립중을 조사하였으며 농업과학기술 연구조사분석기준(RDA, 2012)에 따라 수행되었다.
Total RNA 추출 및 Quantitation Real-Time PCR 분석
포트 시험에서 출수 전 15일에 몰리브덴(MoO42-) 비료를 처리하였으나 포기 내 출수기 변이 차이가 커 RNA 발현 분석은 출수 전 17일에 처리한 시료를 사용하였다. 잎은 비료 처리 9시간 후에 채취하였고 이삭은 출수 후 적산온도가 1,000℃일 때 채취하여 액체 질소로 얼려 마쇄하였다. RNA 추출은 분쇄시료를 0.1 g씩 정량한 후 Plant RNA extraction kit (Sigma Co.)를 사용하였다(Sambrook et al., 1989). Turbo Dnase (Invitrogen)을 사용하여 DNA를 제거했고 RNA정제는 RNA clean-up kit (MACHEREY-NAGEL Co.)를 사용하여 제조사의 매뉴얼에 따라 실시했다. cDNA합성은 1 µg의 RNA를 iscript cDNA synthesis kit (Bio-Rad Co.)를 사용하여 제조사의 프로토콜에 따랐고, 합성된 cDNA 6 µl를 주형으로 하여 상보적 primer (Table 1) 1 µl를 SYBR Green Realtime PCR Master Mix (Toyobo) 10 µl와 혼합하여 Quantitation Real-Time PCR을 수행하였다. 각 유전자의 Ct값은 reference 유전자인 ubiquitin의 Ct값으로 보정한 다음 비교하였다.
Table 1.
Primers used for gene expression analysis related to ABA metabolism.
통계처리
통계처리는 R studio (version 4.4.1)를 이용하여 분산분석한 뒤, agricolae library를 사용하여 P<0.05 수준에서 Duncan 다중검정을 통해 유의성을 분석하였다.
결과 및 고찰
몰리브덴 처리 시기에 따른 수발아 경감 효과
포트시험에서 몰리브덴(MoO42-, 이하 Mo로 표기)의 효과적인 처리시기를 규명하기 위해 Mo 수용액을 출수 전 15일과 출수 후 20일에 처리하였다. 출수 전 15일에 Mo 500, 1,000 ppm을 처리한 결과 수발아율은 무처리에 비해 각각 33.8%, 38.1% 감소하였다. 출수 후 20일 Mo을 처리한 결과 수발아율은 Mo 500 ppm에서만 무처리보다 26.0% 감소했고, Mo 1,000 ppm에서는 유의한 차이가 없었다. 본 시험에서 몰리브덴은 출수 이전에 처리한 것이 출수 후에 처리한 것보다 수발아를 경감시키는 것으로 나타났다(Fig. 1).
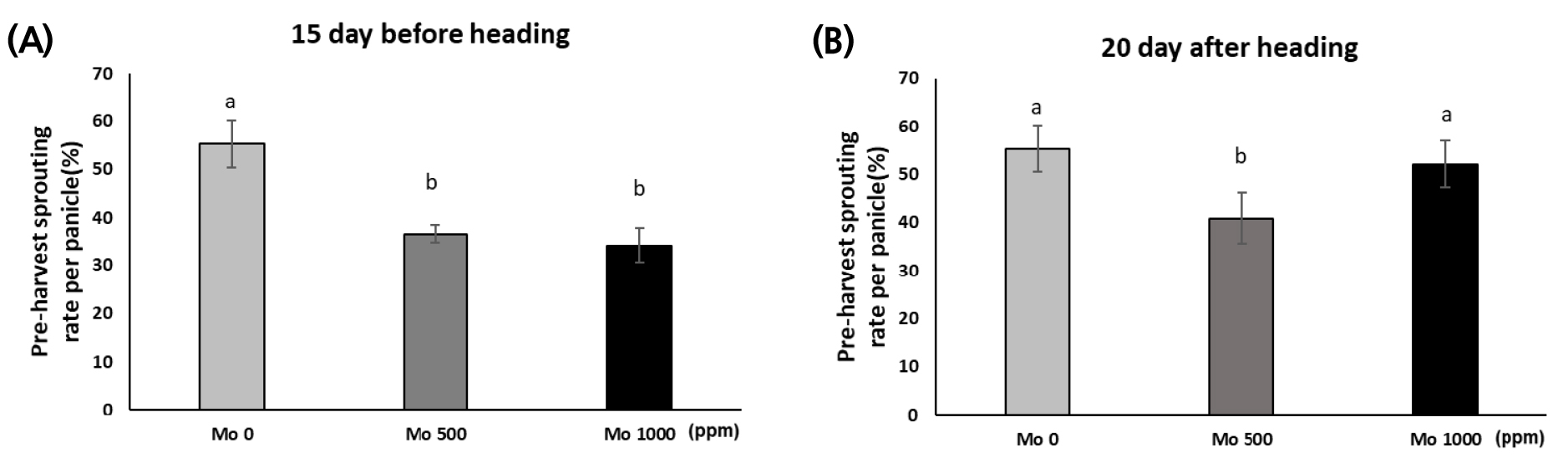
Fig. 1.
Changes in pre-harvest sprouting of Baromi 2 according to the timing of molybdenum solution application (A) Molybdenum treatment at 15 days before heading, and (B) molybdenum treatment at 20 days after heading under water spraying treatment at 22 ± 5°C for four days. Vertical bars represent mean ± S.D. (p < 0.05, n = 3).
몰리브덴 비료 처리량에 따른 수발아 영향
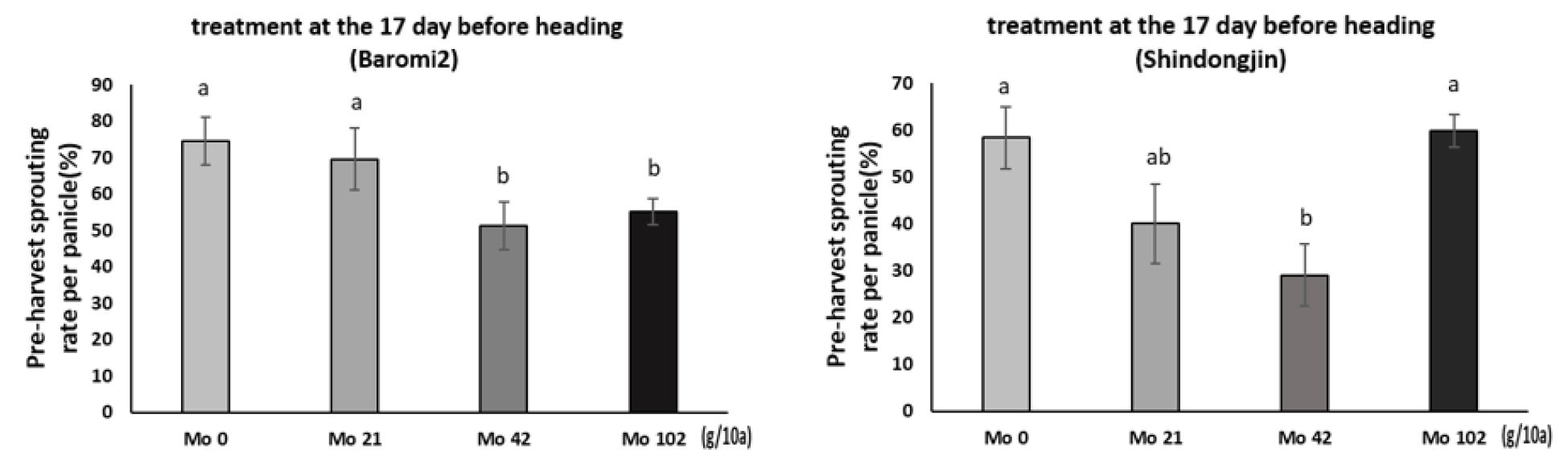
앞서 출수 전에 Mo을 엽면시비하여 수발아가 경감되었음을 확인하고, 액상보다 처리가 간편한 입제 형태로 Mo 비료를 제작하였다. 효과적인 처리시기가 유수형성기이므로 기존에 이삭거름으로 시용되고 있는 NPK복합비료에 몰리브덴산염을 혼합하여 제작하였다. 포트시험에서 유수가 8 cm 신장했을 때를 출수 전 15일로 가정하고 Mo을 처리하였으나 출수기의 변이가 커 바로미 품종은 처리 후 출수까지 12~17일, 신동진 품종은 11~18일 뒤 이삭이 출현하였다(Fig. 2). 몰리브덴 처리 후 출수일까지 기간별로 출수 후 적산온도 1,000℃의 이삭을 채취하여 수발아율을 검정한 결과는 다음과 같다. 분무처리로 수발아를 검정한 결과, Mo 42 g/10a에서 무처리 대비 수발아율은 바로미2는 17.4%, 신동진은 10.8% 감소하였다(Figs. 3, 4A). 벼에서 Mo 100, 600 ppm을 경엽처리시 무처리 대비 수발아율이 유의하게 감소했지만 이보다 고농도인 Mo 3,000 ppm은 효과가 없었다는 보고(Tejakhod & Hammond, 2018)가 있는데 본 시험에서도 가장 높은 처리량인 Mo 102 g/10a에서 수발아율은 무처리와 차이가 없어 Tejakohd (2018)의 보고와 비슷한 경향이었다. 항온항습 조건에서 수발아율을 검정한 결과, 바로미2는 Mo 비료 처리량에 따른 수발아율에 유의한 차이가 없었으나 신동진은 Mo 42, 102g/10a에서 수발아율이 무처리 대비 각각 30.6%, 50.2% 감소하였다(Fig. 4B).
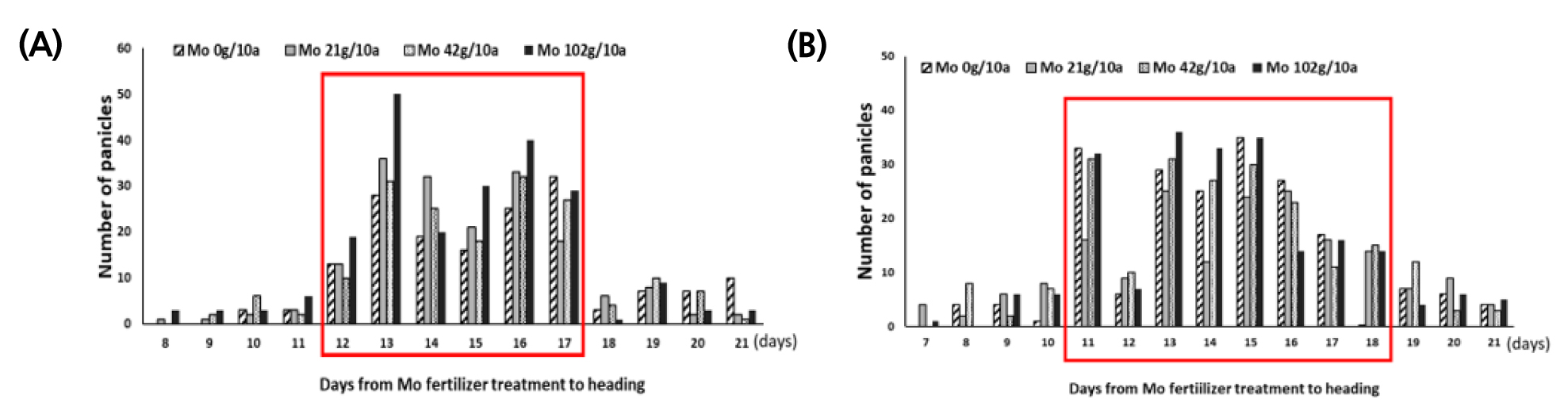

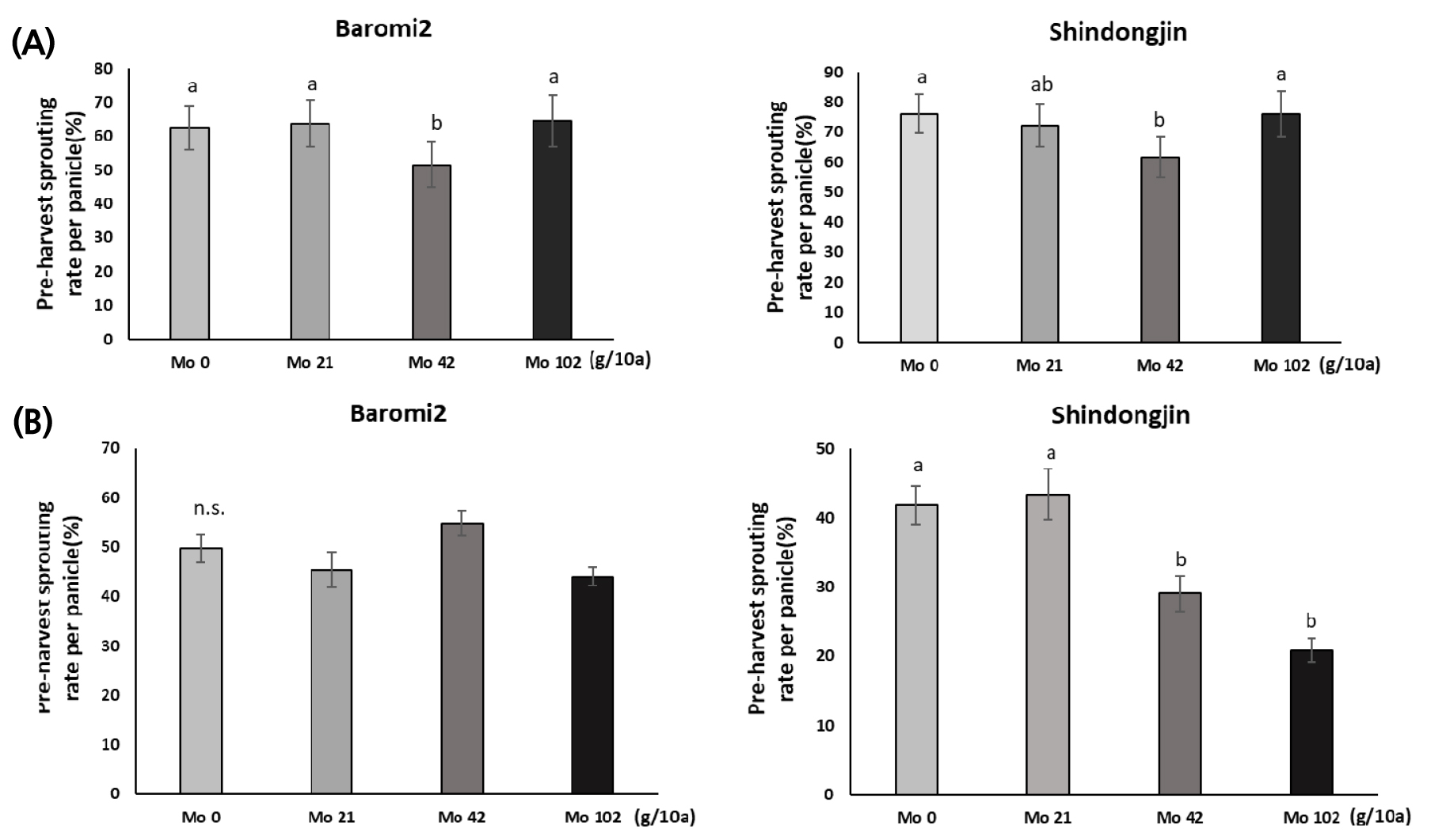
Fig. 4.
Pre-harvest sprouting rate according to molybdenum application amount at 15 days before heading date. (A) Water was sprayed over a duration of four days at 22 ± 5°C. (B) The saturated humidity treatment was conducted over a duration of four days at 25°C. Vertical bars represent mean ± S.E. (p < 0.05, n = 60).
Mo 처리일로부터 이삭별로 출수일의 변이가 있어 이삭별(몰리브덴 처리일~출수일)로 분석한 결과, 수발아 검정 방법 중 분무처리는 두 품종 모두 몰리브덴 처리에 따른 수발아 변화 양상이 비슷했지만 항온항습 조건은 품종 별로 수발아 변화 정도가 달랐다. 그러나 두 방법 모두 무처리 대비 Mo 42g/10a에서 수발아율이 감소하였다. 분무처리 후 시기별로 수발아율을 분석한 결과, 두 품종 모두 출수 전 17일 처리에서 수발아율이 크게 감소하였다. 바로미2의 수발아율은 무처리 대비 Mo 42, 102g/10a 처리구에서 각각 31.1%, 25.9% 감소했고 신동진은 Mo 42 g/10a처리구에서 무처리 대비 50.3% 수발아율이 감소하였다(Fig. 5A). 항온항습 조건은 바로미2는 처리량별로 수발아율의 변화가 없었으며 신동진은 처리 농도가 높을수록 수발아율이 감소하는 경향이 있었다(Fig. 5B).
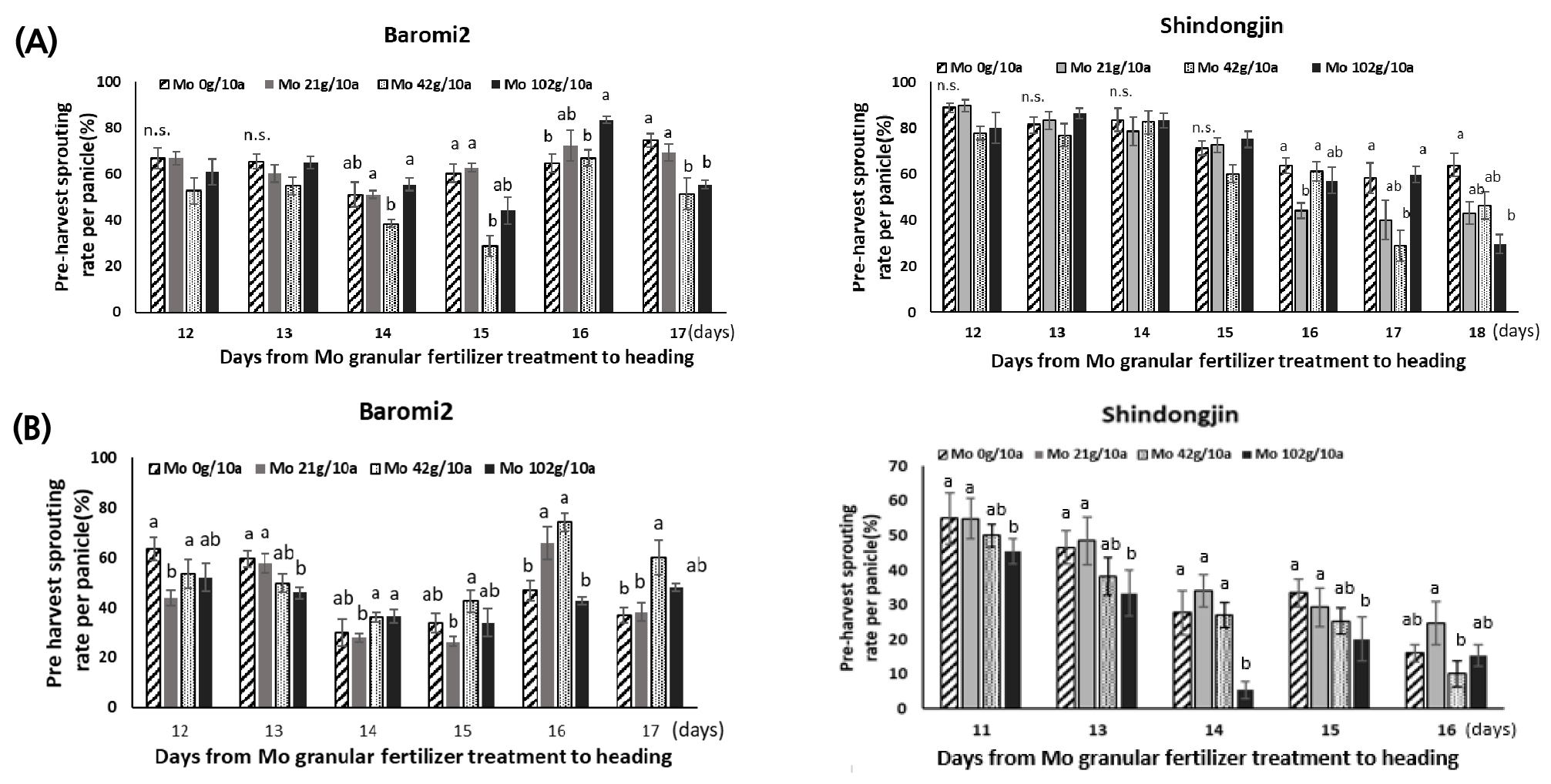
Fig. 5.
Pre-harvest sprouting rate according to the duration from molybdenum fertilizer treatment before heading to the heading date. (A) Water was sprayed over a duration of four days at 22 ± 5°C. (B) The saturated humidity treatment was conducted over a duration of four days at 25°C. Vertical bars represent mean ± S.E. (p < 0.05, n = 10).
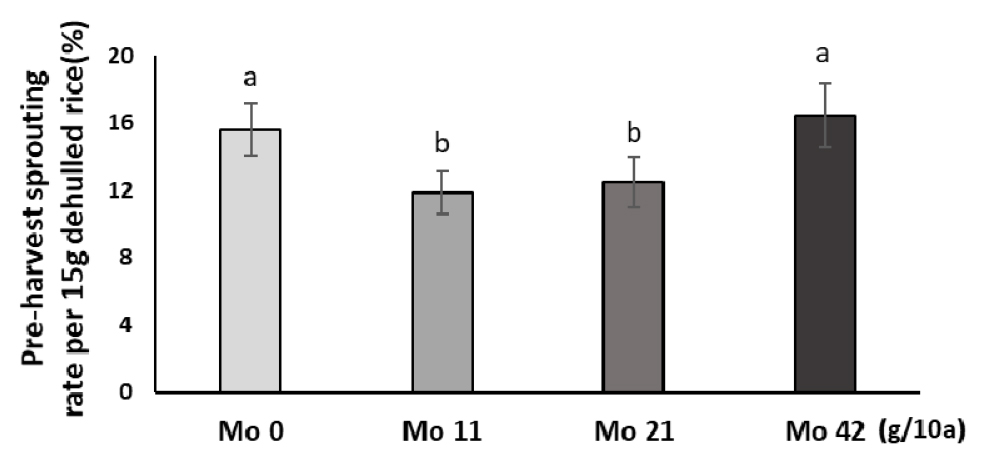
포트시험에서 Mo 처리가 수발아 경감 효과가 있음을 확인하였고, 포장에서 Mo비료 효과를 분석하였다. 적산온도 1,200℃에서 수발아율을 분석한 결과, 무처리 대비 Mo 11, 21 g/10a에서 각각 23.9%, 20.1% 감소하였으나 Mo 42 g/10a에서 수발아율이 증가하여 무처리와 비슷했다(Fig. 6). 이와 같이 포장시험에서도 몰리브덴 처리에 따른 수발아 억제 효과를 확인하였는데 포트시험 결과와 비슷하게 가장 높은 처리량에서 수발아율이 다시 증가했다. 이와 같은 결과로 보아 몰리브덴은 일정 농도에서 수발아 경감에 효과가 있으며 일정 농도 이하와 고농도에서는 오히려 수발아 억제효과가 감소하는 것으로 보인다. 그러나 포트 및 포장 시험에서 수발아 경감에 가장 효과적인 처리량이 달라 추가적인 시험을 통해 면밀한 분석이 필요하다.
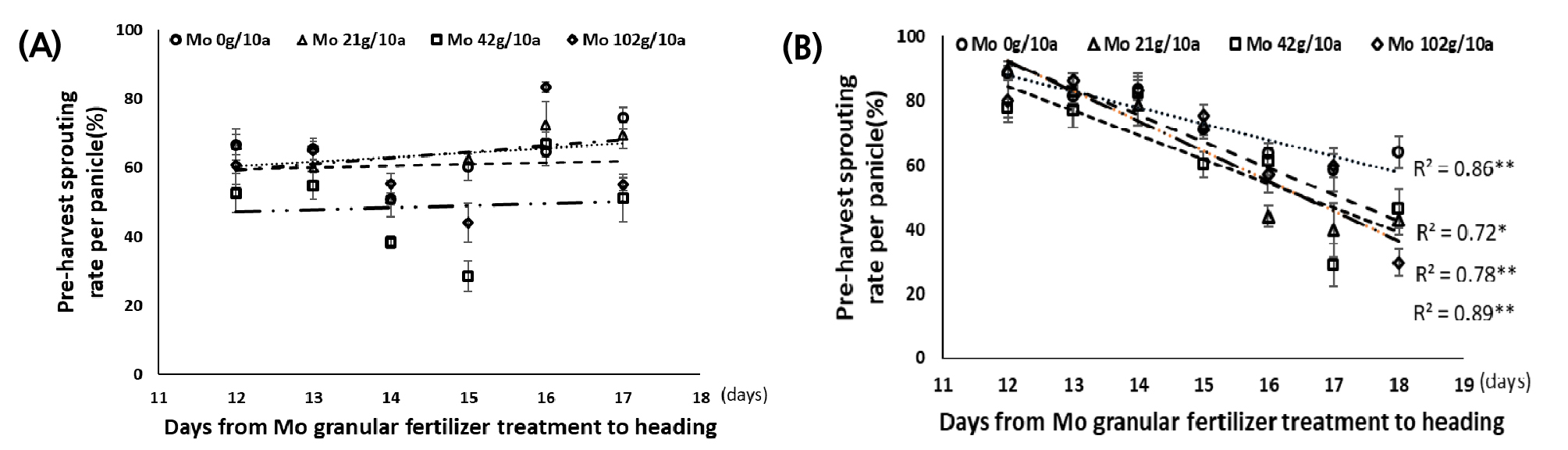
비료처리일에서 출수기까지 경과일수와 몰리브덴 농도별 수발아 차이
포트시험에서 비료처리일부터 출수기까지 경과일수와 처리농도별 수발아율 간에는 품종별로 경향이 달랐다. 분무처리 조건에서, 바로미2는 처리일수와 수발아율 간의 낮은 상관관계를 나타냈다(Fig. 7A). 반면 신동진은 비료 처리량별 비료처리일에서 출수기까지 일수와 수발아율 간의 높은 부의 상관관계를 나타내었다(Fig. 7B). 몰리브덴 처리 후 출수기까지 경과일수별 수발아율을 보면 신동진은 경과일수가 길수록 수발아율이 감소했으며 수발아 감소율은 무처리구에 비해 몰리브덴 처리구의 감소율이 더 큰 것으로 나타났다. 무처리구는 처리일수가 1일 증가할 때마다 5.03% 감소했고 Mo 21 g/10a 처리구에서 9.32%, Mo 42 g/10a 처리구에서 7.55%, Mo 102 g/10a 처리구에서 8.21%로 일별 수발아율이 감소했다(Fig. 7B).
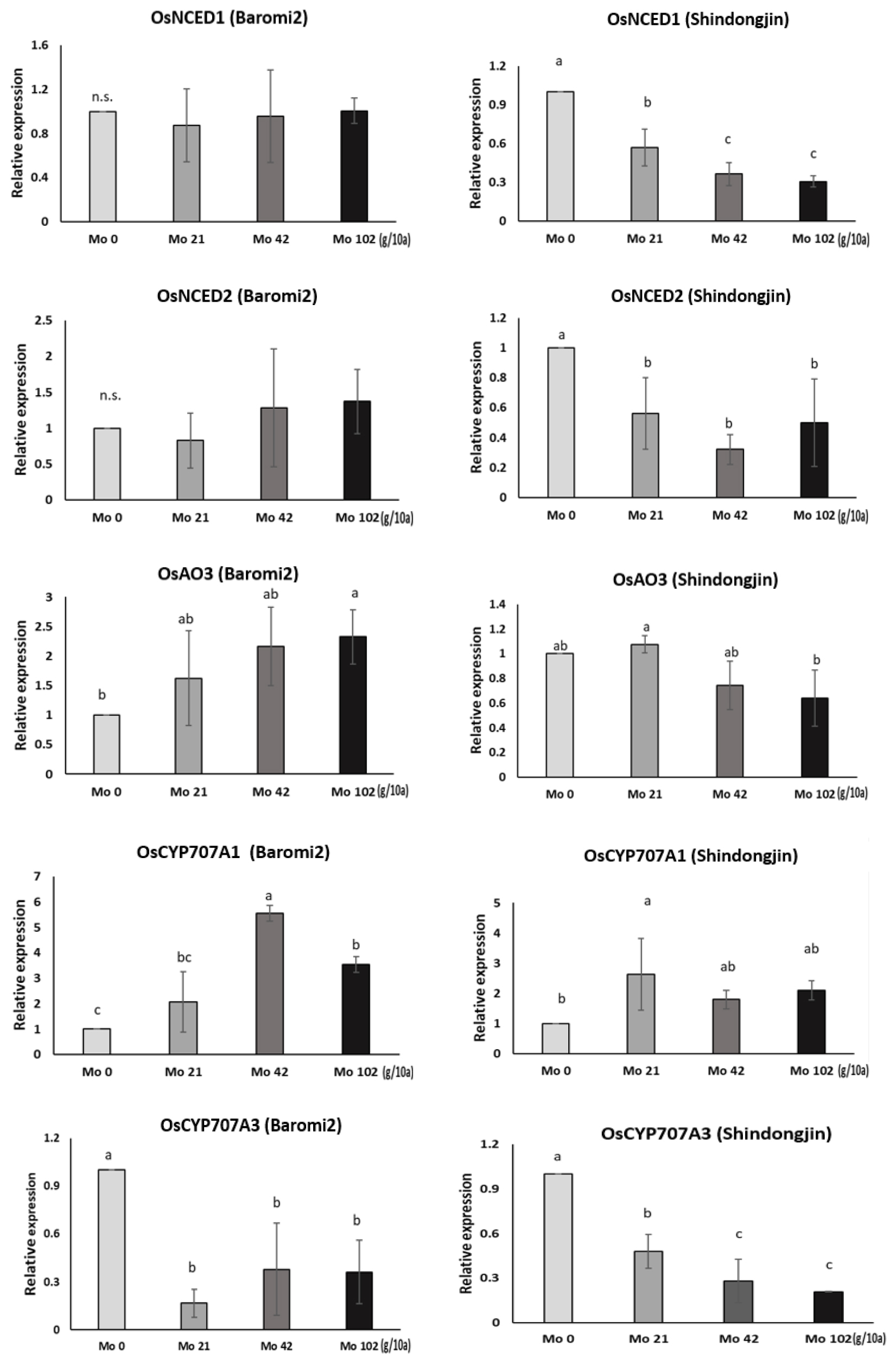
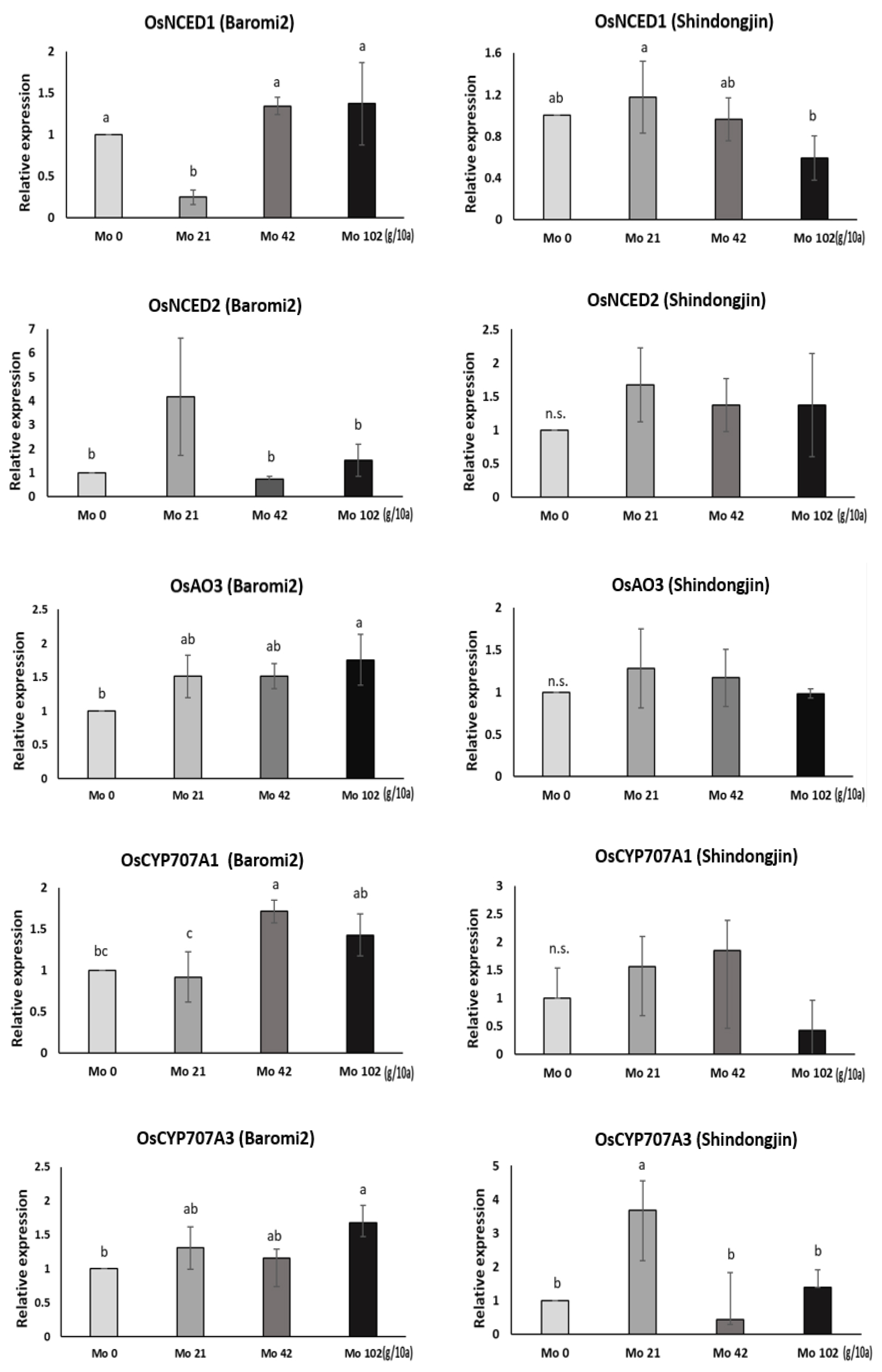
몰리브덴 처리에 따른 ABA 대사 관련 유전자 발현 분석
포트시험에서 Mo 비료처리일부터 출수기까지 경과일수 별로 수발아율을 분석한 결과, 출수 전 17일에 Mo을 처리한 개체의 수발아율이 가장 크게 감소하는 경향을 보였다(Fig. 8). 따라서 이 시기에 Mo 비료 처리 9시간 후의 최상위 3개의 잎과 출수 후 적산온도 1,000℃의 이삭을 채취하여 ABA대사 관련 일부 유전자 발현을 분석하였다. ABA 생합성 과정의 유전자는 OsNCEDs (Oryza sativa 9-cis-epoxycarotenoid dioxygenase), OsAO3 (Oryza sativa Aldehyde Oxidase 3), OsCYP707A (Oryza sativa Cytochrome P450)를 선택하여 quantitation real-time PCR분석을 통해 발현량을 분석하였다. OsNCED 유전자는 ABA생합성 초기에 관여하고 OsAO3 유전자는 ABA생합성 후기에 관여하며, OsCYP707A는 ABA 분해 역할을 하는 것으로 알려져 있다(Zhu et al., 2009). 특히 Aldehyde Oxidase 3 (AO3)는 ABA생합성 과정의 최종단계를 촉진하는데, 몰리브덴을 주성분으로 하는 조효소가 AO3의 활성을 돕는다. 따라서 몰리브덴 처리시 몰리브덴 조효소(MoCo)의 활성으로 AO3의 합성을 촉진하여 ABA함량이 증가된다고 보고되었다(Mendel, 2013; Zhang et al., 2016; Watanabe et al., 2018).
몰리브덴 처리 후 9시간 뒤 최상위 3개 잎의 유전자 발현량을 보면 바로미2의 OsNCED1은 처리량 별로 변화가 없는 반면, 신동진은 처리량이 증가할수록 발현량이 감소하는 경향이 있다(Fig. 9). OsNCED2는 바로미2에서 처리량이 증가할수록 증가하는 경향을 보였다(통계적 유의성은 없었음). 신동진은 무처리구에 비해 처리구에서 낮은 발현량을 보였다. OsAO3의 발현량은 바로미2에서 처리량이 증가할수록 증가하는 경향을 보였고 신동진은 증가하다가 42 g, 102 g/10a에서 무처리 대비 발현량이 감소했다. ABA 분해에 관여하는 유전자인 OsCYP707A1에서 신동진과 바로미 모두 무처리 대비 처리구의 발현량은 높은 반면 OsCYP707A3는 무처리구 대비 처리구의 발현량이 낮았다. 바로미2는 잎에서 몰리브덴 처리 후 ABA 생합성 후반부에 관여하는 OsAO3 유전자의 발현량이 처리량에 따라 증가하였지만 신동진은 처리량에 따라 OsNCED1, OsAO3 발현량이 감소했다. 이처럼 품종별로 발현 양상이 달라 ABA함량 측정을 통해 유전자 발현량과 비교 분석이 필요할 것으로 보인다.
이삭은 등숙적산온도 1,000℃에 채취하여 조사하였다. 출수 전 17일에 Mo을 처리한 이삭의 수발아율을 보면 바로미2는 처리량이 증가할수록 수발아율이 감소하는데(Fig. 8), ABA생합성 초기와 후기에 관여하는 유전자 발현량이 증가하여 수발아에 영향을 준 것으로 보인다. OsNECD1 유전자에서 무처리 대비 42 g, 102 g/10a에서 약간 증가했고 21 g/10a구간은 발현량이 무처리보다 낮았지만 OsNECD2에서 21 g/10a발현량이 가장 높았다. OsAO3는 처리량이 높아질수록 발현량이 증가했다. 그러나 몰리브덴 처리량이 많아질수록 ABA 분해와 관련된 유전자인 OsCYP707A1,2의 발현량도 높아졌다(Fig. 10). 신동진 이삭의 수발아율을 보면 몰리브덴 처리량이 증가할수록 감소하다가 급격히 높아지는 구간이 나타났다(Fig. 8). Mo 42g/10a에서 가장 수발아율이 낮았는데 통계적으로 유의하지는 않았지만 무처리 대비 상대적으로 OsNCED2와 OsAO3의 발현량이 높았다. 수발아율이 가장 높았던 Mo 102g/10a구간은 OsNCED1 발현량이 무처리구에 비해 현저히 낮았고 OsAO3 발현량도 무처리와 비슷한 수준이었다.
종합적으로 바로미2의 종자와 잎에서 OsNCED, OsAO3 발현량을 보면 무처리 대비 몰리브덴 처리구에서 증가했고 수발아율은 감소한 것을 확인할 수 있었다(Figs. 9, 10). 하지만 분해유전자인 OsCYP707A1,2의 발현량도 증가했는데 세포 내 ABA 수준은 호르몬의 생합성(biosynthesis)과 분해 간의 균형에 의해 결정된다는 보고처럼 ABA 생합성 관련 유전자의 영향이 상대적으로 커 수발아율이 감소한 것으로 생각된다(Nambara et al., 2005). 신동진은 처리량이 증가할수록 수발아가 감소하다가 Mo 102 g/10a 처리구에서 증가했는데, 종자의 유전자를 분석한 결과 ABA생합성, 분해 관련 유전자의 발현량이 증가하다가 감소하는 패턴을 보였다(Fig. 10). 이처럼 ABA 생합성 관련 유전자의 발현량이 증가하다 감소하는 구간을 보면 고농도의 몰리브덴은 오히려 수발아 억제 효과가 없는 것으로 보인다. 이때 ABA 분해 관련 유전자도 함께 증가하다가 감소하는 구간이 있어 추후 ABA 함량과 비교 분석이 필요하다.
ABA 생성에 영향을 미쳐 몰리브덴 처리 후 초기에 관찰된 OsAO3 발현량이 등숙 후기에도 몰리브덴 농도별로 비슷한 패턴으로 변화한(Figs. 9, 10) 것으로 보아 출수 이전에 처리한 몰리브덴이 출수 후기까지 ABA 생합성 과정에 관여할 것으로 생각되며, OsNCED와 OsAO3 발현량 조절을 통해 수발아율에 영향을 미치는 것으로 보인다. 다만 품종간 발현양상의 차이 등은 추가적인 생리연구가 필요하다.
몰리브덴 처리에 따른 수량구성요소 변화
포트시험에서는 품종별로 몰리브덴 처리량에 따른 영화수, 등숙률, 천립중의 유의한 변화는 없었다(Table 2). 영화수는 바로미2에서 몰리브덴 처리량이 증가할수록 증가되는 경향을 보였으나 통계적 유의성은 없었다. 포장실험은 Mo 11 g/10a에서 정조 수량이 가장 높았는데 면적당 이삭수가 많아져 수량이 증가한 것으로 보인다. 현미 수량 및 수량구성요소들은 Mo 처리량별로 유의한 차이는 없었다(Table 3). 벼에서 Mo 무처리, 100, 600, 1,000 ppm 처리시 천립중은 비슷했지만 100, 600 ppm 처리시 무처리 대비 수량이 증가했다는 보고가 있다(Tejakhod & Hammond, 2018). 콩과 옥수수에서 몰리브덴을 30 g/ha 농도로 엽면 시비 시 질산 환원 효소(NR) 활성이 증가했고 잎의 질소 함량과 단백질 합성을 촉진함으로써 작물의 수확량을 증가했다는 연구 결과가 있어 (Oliveira et al., 2022) 추후 포장시험에서 몰리브덴 처리가 수량에 영향을 미치는지 면밀히 분석할 필요가 있다.
Table 2.
Differences in rice yield components of Baromi2 and Sindongjin under molybdenum treatment in the pot experiment. Data are mean ± S.D. (p < 0.05, n = 4).
Table 3.
Differences in rice yield and yield components of Baromi2 under molybdenum treatment under field conditions. Data are mean ± S.D. (p < 0.05, n = 4).
적 요
본 연구는 몰리브덴이 바로미2와 신동진의 수발아를 경감하는 효과 및 적정 처리시기와 처리농도를 구명하고자 포트시험과 포장시험을 통해 확인하였으며 그 결과를 요약하면 다음과 같다.
1.포트시험은 몰리브덴 수용액을 출수 전 15일과 출수 후 20일에 처리하였고 출수 이전 처리가 출수 후 처리보다 수발아 경감에 효과있는 것으로 나타났다.
2.포트시험 결과, 출수 전 15일 Mo 처리 시 분무처리를 통한 수발아검정 방법에서는 바로미2, 신동진 모두 Mo 42 g/10a 처리구에서 무처리 대비 수발아율이 감소하였다. 반면 항온항습 조건에서 수발아를 검정한 경우, 바로미2는 처리량별 차이가 없었지만 신동진은 Mo 42 g, 102 g/10a에서 수발아율이 감소하였다. 포장실험에서 출수 전 25일에 Mo을 처리한 결과 바로미2는 Mo 11, 21 g/10a에서 유의하게 수발아율이 감소하였으나 Mo 42 g/10a에서는 오히려 수발아율이 증가하였다.
3.포트시험에서, 분무처리를 한 바로미2는 비료 처리일에서 출수기까지 경과일수와 수발아율 간의 낮은 상관관계를 보였지만 신동진은 비료 처리량별로 비료 처리일에서 출수기까지 경과일수와 수발아율 간의 높은 부의 상관관계를 나타냈다. 또한 무처리 대비 몰리브덴 처리구의 일별 수발아 감소율이 더 컸다.
4.포트시험에서, 출수 전 17일에 몰리브덴을 처리한 이삭과 잎을 분석한 결과, 몰리브덴 처리량이 높을수록 바로미2에서 수발아율은 낮아졌는데 ABA 합성 관련 유전자 및 분해 관련 유전자의 발현량이 높아지는 경향을 보였다. 그러나 신동진은 몰리브덴 처리량에 따라 수발아율이 감소하다가 증가하는 구간이 있었고 ABA 생합성 관련 유전자의 발현량도 증가하다가 감소하는 구간이 있어 고농도의 몰리브덴에서는 수발아 억제 효과가 없는 것으로 생각된다.
5.포트시험과 포장시험 모두 몰리브덴 비료 처리량에 따른 영화수, 등숙률, 천립중의 유의한 변화는 없었으나, 포장시험에서 수량은 Mo 11 g/10a 처리 시 유의하게 증가했다. 다만, 포장시험은 1년차 시험 결과로 추후 검증이 필요할 것으로 보인다.
