

서 론
재료 및 방법
시험재료 및 생육상 조건
뿌리 및 지상부 생육량 조사
엽록소 함량
항산화 활성 및 성분 분석
통계분석
결과 및 고찰
근권 생육 온도에 따른 뿌리 및 지상부 생육 특성
생육온도 차이에 따른 고구마 잎의 엽록소 함량 변화
생육온도 차이에 따른 품종별 항산화 활성 및 2차 대사물질의 함량 변화
적 요
서 론
고구마는 열대 및 아열대 지역의 고온성 뿌리작물로 저온에 매우 민감하다(Fujiwara et al., 2004; Osaki et al., 1996; Wees et al., 2016). 저온 피해는 발생 온도와 지속 시간, 작물의 생육 단계, 재배 품종의 특성, 그리고 재배지의 지형과 토양 조건에 따라 다양하게 나타날 수 있다(Choi and Lee, 1976). 고구마는 토양 온도 25℃인 조건에서 덩이뿌리의 형성이 가장 잘 이루어지며, 토양 온도가 낮은 15℃ 조건에서는 뿌리 발생 및 비대가 억제된다고 보고하였다(Ueki & Sasaki, 1987). 또한 Eguchi et al. (2003)은 16℃의 낮은 온도 조건에서 fibrous roots(섬유근)의 생장이 현저하게 억제된다고 보고한 결과로 볼 때 15~16℃는 고구마의 성장을 억제하는 하한 온도로 간주되며, 온도는 괴근 분화를 포함하여 고구마의 괴근 비대 속도, 줄기 신장, 잎 면적 확장 및 등 여러 성장 및 발달 과정에 영향을 미친다고 하였다(Gajanayake et al., 2014). Wijewardana et al. (2018)은 고구마 정식 후 초기에 주간 22/야간 14℃의 낮은 온도에서는 최적 온도 30/22℃에 비해 넝쿨 길이 95%, 마디 수 70%, 잎 면적 78%, 총 바이오매스 66% 및 순 광합성량 36%로 현저하게 감소한다고 하였다. 정상적인 조건하에서 괴근 분화 개시는 정식 후 2주 후부터 시작되기 때문에 정식 후 처음 14일에서 30일 동안이 식물당 분화된 괴근 수와 최종 수확량에 직접적인 영향을 미친다고 강조하였다(Meyers et al., 2017; Villordon et al., 2012). Wrigley et al. (1994)은 온도 스트레스는 작물 성장과 발달에 가장 결정적인 제한 중 하나로 최종 수확량에 영향을 미친다고 하였다. 최근 5년간 우리나라의 고구마 재배면적은 18.4~23.2천 ha로 생산량은 30~36만 톤 수준으로 밭작물 중에서 ha 당 소득이 17.8~13.4백만 원으로 가장 높은 작물이다(AFRASY, 2023). 고구마 재배에 적합한 계절은 5월 상순에서 10월 하순까지이나 남부지역에서는 4월 상순부터 정식하여 8월 상순에 수확하거나 재배기간을 150일 이상 늘려 수확하는 재배면적이 증가하는 추세이다. 정식 후 생육 초기인 4월의 낮은 기온과 예상되지 않은 서리 내림으로 식물체 고사 등 저온 피해로 인해 재 정식, 생육 불량 및 수량 감소 등의 원인이 되고 있다. 따라서 본 연구는 고구마 정식 후 생육 초기에 온도 차이에 따른 뿌리, 줄기, 잎 생장량, 그리고 엽록소 함량, 생리활성 반응 등을 분석하여 생육초기 저온이 고구마 묘 생육에 미치는 기초자료를 얻고자 수행하였다.
재료 및 방법
시험재료 및 생육상 조건
본 연구에 사용된 시험재료는 전남 무안 소재의 국립식량과학원 바오에너지작물연구소 내 고구마 묘 생산을 위한 연동 하우스 시설내의 묘상에 2024년 2월 10일에 씨고구마를 파종하여 생육시킨 묘를 재료로 하였다. 품종은 최근 육성된 육색이 황색인 ‘소담미’, 연황색의 ‘진율미’, 그리고 주황색인 ‘호풍미’를 사용하였다. 묘 조건은 3품종 동일하게 묘의 끝순 마디에서 아래쪽 6마디부터 8마디에 해당하는 3마디를 자르고 잎을 제거하였다. 생육상(다솔과학, DS-95L)의 일장 주기는 주간 14시간, 야간 10시간으로 고정하여 주간에 광량은 1000 lux 수준으로 유지하였다. 온도는 주간/야간 17/13, 19/15, 그리고 21/17±0.5℃로 3대의 생장상에서 동시에 실험하였다. 묘의 정식은 스티로폼 배드(가로-세로 높이)에 시판용 모래(에코샌드)를 채우고 품종별로 준비된 묘를 3마디 중 2마디가 묻히도록 비스틈하게 50주씩 정식하여 생육시켰다. 스프로폼 배드 내 토양 근권 온도는 토양 온습도 로거(에펜토, SM)을 이용하여 모니터링 하였다.
뿌리 및 지상부 생육량 조사
생육상 온도 조건별 뿌리 수 및 무게, 엽수, 엽면적, 줄기신장 그리고 지상부 무게는 정식 후 7일부터 7일 간격으로 35일까지 5회 조사하였다. 조사 개체는 온도 및 품종별 각각 10주씩 뿌리가 떨어지지 않게 캐내어 흐르는 물에서 모래를 씻어낸 다음 물기를 제거하고 개체별로 조사하였다. 엽면적은 LI-3100 Area Meter (LI, COR, inc, Lincoln, Nebraska. USA)로 측정하였다.
엽록소 함량
고구마 잎의 동일 부위에서 엽록소를 추출하기 위해 직경 5 mm의 코르크 보러(cork borer)를 사용하여 엽면적을 채취하였다. 약 200 mg의 준비된 엽 조직을 E-tube에 넣고, 95% 에탄올(EtOH) 1 mL를 첨가하였다. 이를 80℃의 항온 수조에서 20분간 가열하여 엽록소를 추출하였다. 가열 중 10분 경과 시점에서 튜브를 흔들어 혼합물을 균일하게 섞어주었다. 가열이 완료된 시료는 즉시 얼음 위에서 냉각시켜 추가적인 변화를 방지하였다. 냉각된 추출물을 사용하여 분광광도계로 664 nm 및 648 nm에서의 흡광도를 측정하였다. 이때, 95% EtOH를 blank로 사용하여 흡광도를 보정하였다. 엽록소 A와 B의 함량은 다음 식을 사용하여 계산하였다:
엽록소 총함량은 엽록소 A와 B의 농도를 각각 산출하여 더하였다.
항산화 활성 및 성분 분석
고구마 지상부 조직을 수확하여 동결건조기(FDTA-5050, Operon)로 건조한 후 분쇄하여 분석에 사용하였다. 총 폴리페놀 분석은 동결 건조된 시료 0.25 g의 methanol 추출물 100 μL에 2% Na2CO3 용액 2mL를 가한 후 3분간 방치하여 50% Folin-Ciocalteu reagent (Sigma-Aldrich, St. Louis, MO, USA) 100 μL를 가한 후 3,000 rpm에서 10분간 원심 분리하여 실온에서 반응시켰다. 반응액은 UV/Vis spectrophotometer를 사용하여 750 nm에서 흡광도 값을 측정하였다. 표준물질로 gallic acid (Sigma-Aldrich, St. Louis, MO, USA)를 사용하였으며, 검량선을 작성하여 총 폴리페놀 함량은 시료 100 g 중의 mg garlic acid equivalent (dry basis)로 나타낸 후 대조구 대비 %로 표시하였다.
총 플라보노이드 함량은 시료 추출물 추출물 250 μL에 증류수 1 mL와 5% NaNO2 75 μL를 가한 다음, 5분 후 10% AlCl3·6H2O 150 μL를 가하여 6분 방치하고 1N NaOH 500 μL를 가한 후 11분 뒤에 반응액의 흡광도 값을 510 nm에서 측정하였다. 표준물질인 (+)-catechin hydrate(Sigma-Aldrich, St. Louis, MO, USA)를 사용하여 검량선을 작성하였고, 시료 100 g 중의 mg catechin equivalent (dry basis)로 나타낸 후 대조구 대비 %로 표시하였다.
DPPH (1,1-diphenyl-2-picryl hydrazyl, Sigma-Aldrich, 미국) 라디칼 소거 활성은 추출물 200 μL에 0.2 mM DPPH 용액 800 μL 첨가 한 후 10초간 진탕하고 30분 후 525 nm에서 흡광도를 측정하였다. Trolox로 검량선 작성하고, 시료 100 g 중 mg Trolox equivalent antioxidant capacity(TEAC)로 계산 후 대조구 대비 %로 표시하였다.
통계분석
모든 실험 결과는 SPSS 소프트웨어 버전 29 (IBM, Chicago, IL, USA)을 이용하여 p<0.05, p<0.01, p<0.001 수준에서 분산분석(two-way ANOVA)과 p<0.05 수준에서 Duncan의 다중범위 검정으로 시료 간의 유의성을 검정하였고, 데이터는 10반복의 평균값으로 나타내었다.
결과 및 고찰
근권 생육 온도에 따른 뿌리 및 지상부 생육 특성
조기재배 시 주로 재배 되고 있는 진율미 품종과 새롭게 육성된 두 품종 ‘호풍미’와 ‘소담미’의 저온에 대한 생육 특성을 비교하기 위하여 조기재배 정식 시기인 4월 평균기온의 범주 안에서 주간/야간 17/13, 19/15, 그리고 21/17±0.5℃로 온도를 설정하여 생장상에서 세 품종의 생육 특성을 조사하였다. 근권 토양의 온도를 3일 동안 모니터링한 결과, 21/17℃로 설정한 온도에서는 21.3℃~15.3℃, 19/15℃의 설정 온도는 19℃~14.5℃, 그리고 17/13℃의 설정은 17℃~12.5℃의 범위로 최고 최저 온도를 반복하여 변화하는 것을 확인하였다(Fig. 1). 고구마는 12℃ 미만에서는 생존할 수 없었으며, 15℃ 이상에서 뿌리가 내리고 20℃ 이상에서 정상적인 생육이 이루어진다(Harter & Whitney, 1962). 그러므로, 이 시험에서 설정된 온도 범위는 이러한 고구마 저온 생육 특성을 파악할 수 있는 적정한 온도로 고려되었다. 생육 특성 조사 결과, 세 품종 모두 17/13°C 처리에서의 뿌리 형성은 19/15°C 및 21/17°C 처리와 비교하여 현저하게 감소하였다(Fig. 2). 특히 ‘소담미’의 경우 처리 4주 후 부터 뿌리 수 발생이 현저하게 줄었으며, 5주차에는 77% 감소가 관찰되었다. 반면, ‘호풍미’와 ‘진율미’는 각각 49% 및 58%의 감소를 보였다(Fig. 3). 또한 ‘소담미’는 17/13°C에서 뿌리 무게가 0.03 g으로 가장 낮았고, 21/17°C에서 0.48 g으로 증가하였다. ‘호풍미’와 ‘진율미’ 역시 온도가 증가함에 따라 뿌리 무게가 증가하였으며, ‘호풍미’는 21/17°C에서 0.73 g, ‘진율미’는 0.66 g으로 생장량이 많았다(Table 1). 이는 저온 조건에서 뿌리 형성이 저해되며, 특히 ‘소담미’에서 그 영향이 크게 나타났음을 시사한다. 세 품종의 평균 뿌리 무게를 비교하면, ‘진율미’와 ‘호풍미’가 ‘소담미’보다 상대적으로 더 나은 뿌리 형성 능력을 보였다. 지상부의 생육에서도 유사한 경향이 관찰되었다. 세 품종 모두 21/17°C 조건에서 신초 길이와 신초 및 잎의 생체 중량이 가장 높게 나타났다. 반면에 온도가 낮을수록 특히 17/13°C 조건에서 세 품종 모두 지상부 생육이 저해되었다. 특히, ‘소담미’는 신초 길이가 0.8 cm로 가장 짧았고, 생체 중량도 0.17 g으로 나타나 다른 두 품종에 비해 다소 낮았다 (Table 1). 또한 온도가 낮아짐에 따라 잎 수와 면적이 감소하였으며, 특히 ‘소담미’는 17/13°C에서 잎 수가 1.6개, 잎 면적이 6.8 cm2로 저온에서 가장 저조한 성장을 보였다(Table 1, Fig. 4). 이는 저온 스트레스가 잎의 광합성 활동을 억제 시켜 식물의 생장이 저해된 결과로 보인다.
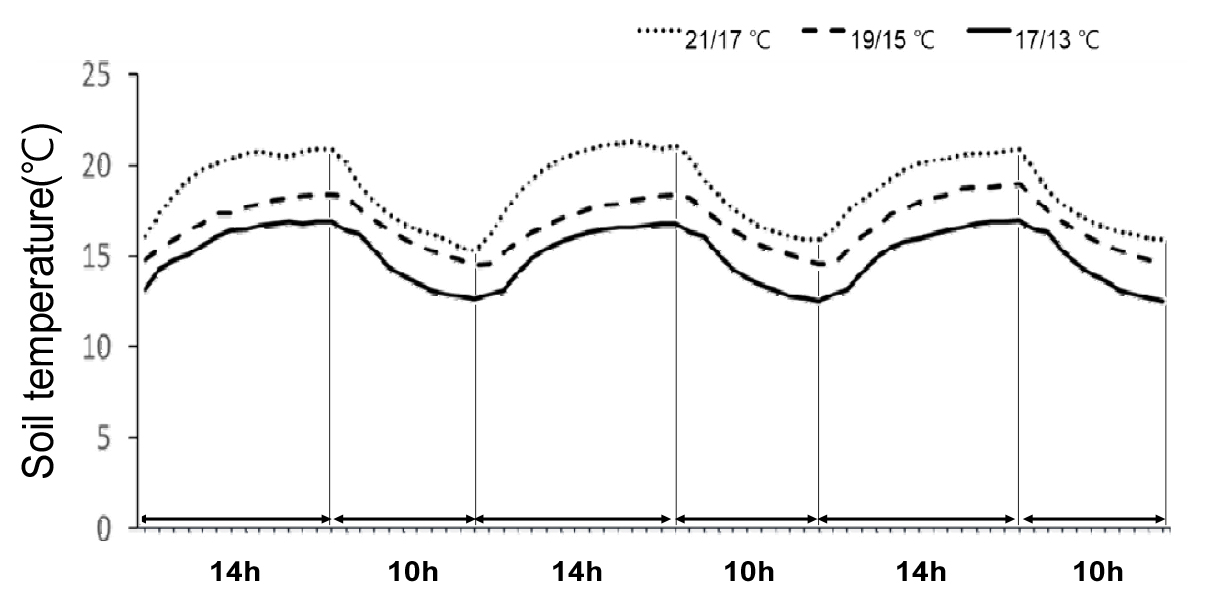
Fig. 1.
Soil temperatures in the pots, monitored for three representative days during the 35-day treatment period. The temperature treatments in the growth chambers were set at three levels: 17/13°C, 19/15°C, and 21/17°C, with the day temperature lasting 14 hours and a night temperature lasting 10 hours.

Table 1.
Root weight, total root length, number of leaves, and leaf area under different growth temperatures at 35 days after sweetpotato cutting.
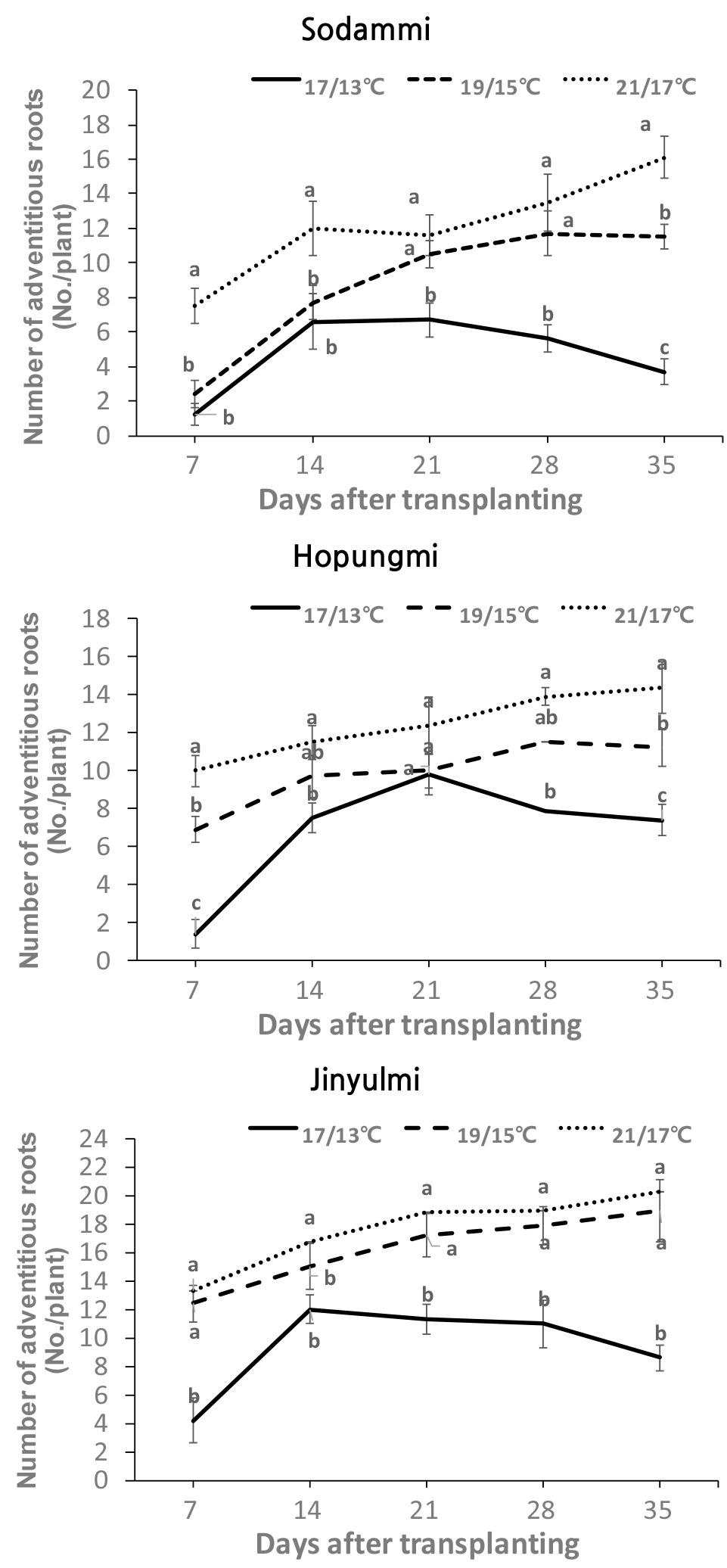
Fig. 3.
Effects of different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) on the number of adventitious roots of the three sweet potato cultivars. Values (mean ± SE) with different letters within the same day indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test.
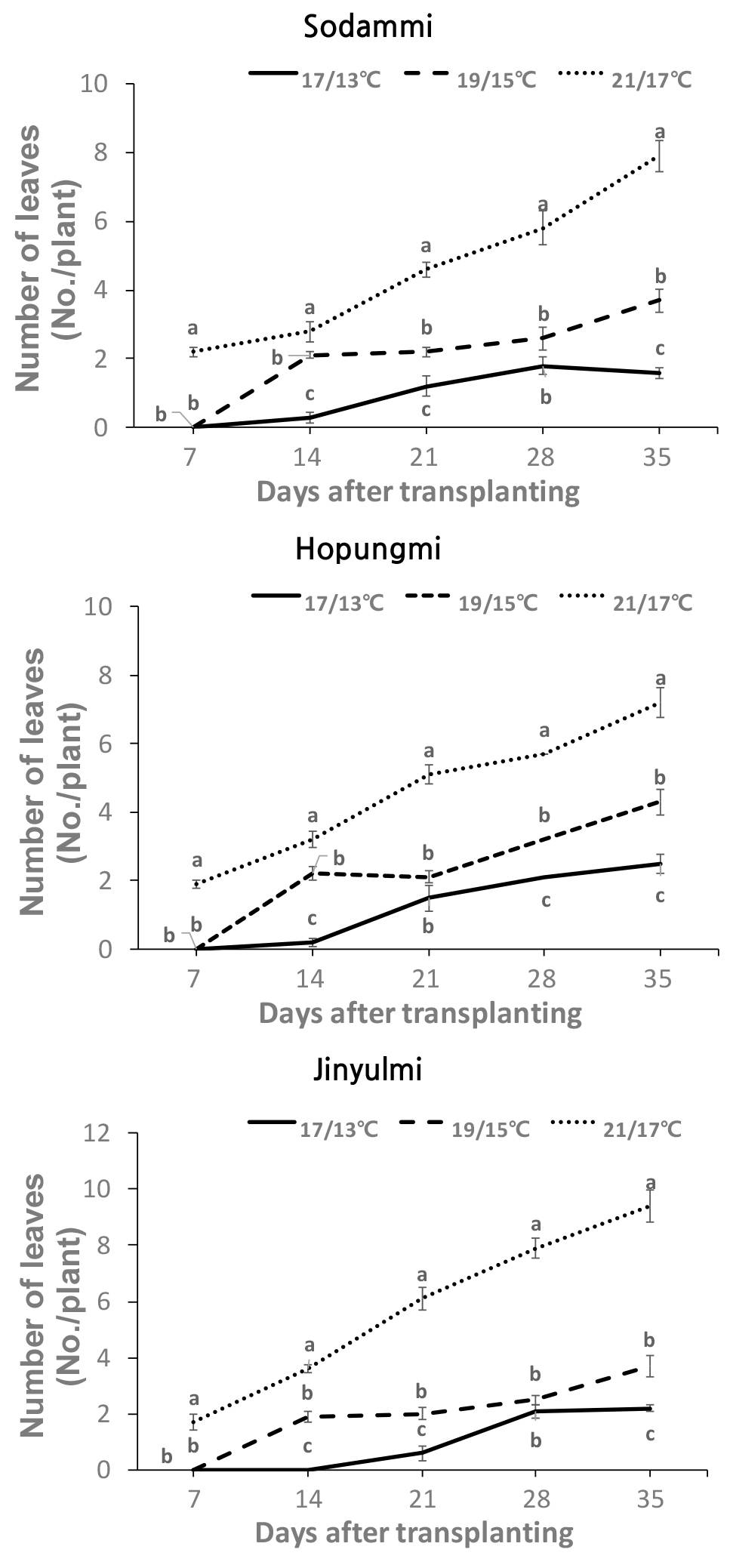
Fig. 4.
Effects of different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) on the number of leaves of the three sweet potato cultivars. Values (mean ± SE) with different letters within the same day indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test.
생육온도 차이에 따른 고구마 잎의 엽록소 함량 변화
처리 5주차에 엽록소 함량(mg/g FW)을 분석한 결과, 처리온도가 낮을수록 엽록소 함량은 감소 하였으며 진율미(49~41), 호풍미(44~32), 소담미(40~25) 순서로 엽록소 함량의 감소 폭이 크게 나타났다(Fig. 5). 저온 스트레스는 식물의 생리적, 생화학적 변화에 중요한 영향을 미치며, 특히 엽록소 함량에 크게 영향을 준다. 본 연구에서 확인된 바와 같이, 처리 온도가 낮아짐에 따라 모든 품종에서 엽록소 함량이 감소하였으며, 이는 고구마 정식 초기에 저온 스트레스는 고구마 광합성 능력을 저하시키는 주요 요인이 될 수 있을 것으로 생각된다(Lin et al., 2007). 저온 환경에서 식물의 엽록소 함량이 감소하는 이유는 엽록소 생합성 과정이 억제되거나 엽록소 분해 과정이 활성화되기 때문이다(Lin et al., 2007). 저온 조건은 엽록체 내 효소 활성을 저하시켜 엽록소 생합성을 방해하며, 동시에 활성산소종(ROS)의 축적을 촉진하여 엽록소 분해를 가속화할 수 있다(Lin et al., 2007). 특히, 본 연구에서는 ‘소담미’에서 가장 큰 엽록소 감소가 관찰되었는데, 이는 해당 품종이 저온 스트레스에 상대적으로 민감하다는 것을 의미한다. ‘호풍미’와 ‘진율미’는 ‘소담미’에 비해 엽록소 감소 폭이 작았으며, 이는 두 품종이 저온 조건에서 상대적으로 더 높은 광합성 능력을 유지할 수 있다는 점을 나타낸다. 따라서, 엽록소 함량의 변화는 저온 스트레스에 대한 식물의 내성 정도를 평가하는 중요한 지표가 될 수 있으며, 엽록소 함량의 유지 능력은 식물의 저온 내성을 높이는 주요 요인으로 작용할 수 있다. 앞으로의 연구에서는 이러한 엽록소 변화와 관련된 유전자 발현 및 대사 경로를 더 면밀히 분석하여 저온 스트레스 내성 품종을 개발하는 데 기여할 수 있을 것이다.
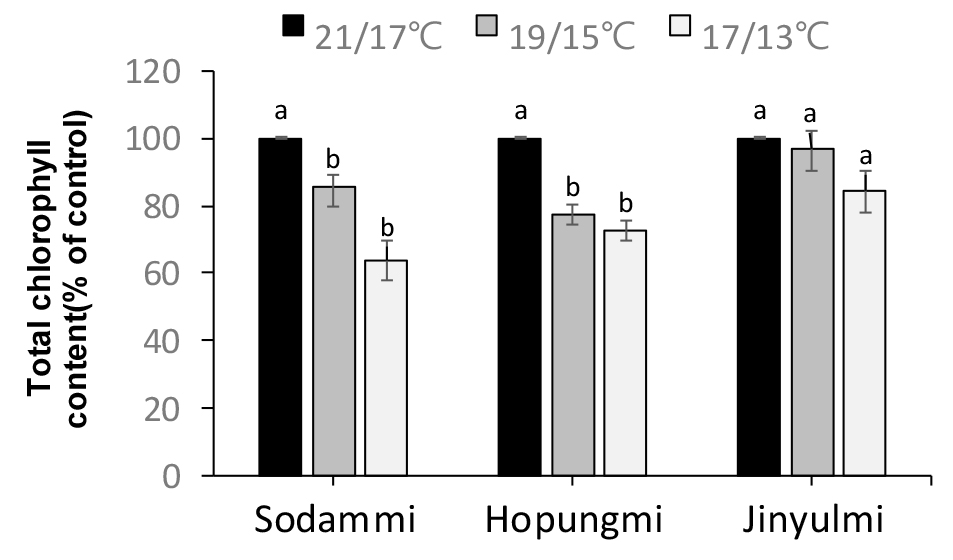
Fig. 5.
Total chlorophyll content in the shoots of three sweetpotato cultivars under different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) for 35 days. Values (mean ± SE) with different letters within the same cultivar indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test. The control refers to the sample cultivated at 21/17°C.
생육온도 차이에 따른 품종별 항산화 활성 및 2차 대사물질의 함량 변화
DPPH 라디칼 소거 활성
DPPH 라디칼 소거 활성(DPPH radical scavenging activity)은 식물의 항산화 능력을 평가하는 주요 지표로, 저온 스트레스와 같은 환경적 스트레스 하에서 활성산소종(ROS)을 제거하는 능력을 반영한다. ‘호풍미’와 ‘진율미’는 17/13°C의 낮은 생육온도에서 DPPH 라디칼 소거 활성이 대조군(21/17°C)에 비해 각각 24%와 7% 증가한 반면, ‘소담미’는 유의미한 변화가 없었다(Fig. 6). 이러한 결과는 저온 스트레스에 대한 품종별 항산화 방어 능력의 차이를 보여주며, ‘호풍미’와 ‘진율미’가 ‘소담미’에 비해 저온 환경에서 더 높은 내성을 지닌다는 것을 시사한다.
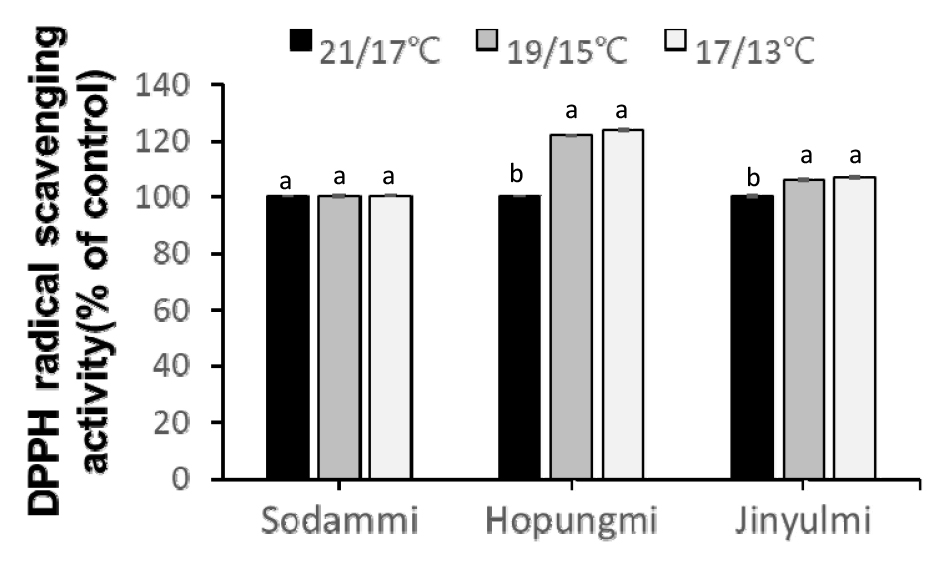
Fig. 6.
DPPH radical scavenging activity in the shoots of three sweetpotato cultivars under different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) for 35 days. Values (mean ± SE) with different letters within the same cultivar indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test. The control refers to the sample cultivated at 21/17°C.
저온 스트레스는 식물에서 ROS의 축적을 유도하여 산화적 스트레스를 증가시키며, 이는 세포막 손상, 단백질 변형, DNA 손상 등의 결과를 초래할 수 있다(Anderson et al., 1988). 식물은 이러한 ROS의 해로운 영향을 완화하기 위해 항산화 효소 및 비효소 항산화제를 동원하여 방어 메커니즘을 강화한다(Salin, 1988). DPPH 라디칼 소거 활성의 증가는 이러한 항산화제들이 저온 스트레스 하에서 활성화되어 ROS를 효과적으로 제거하고, 식물이 산화적 손상을 최소화할 수 있음을 의미한다(Lin et al., 2007). 특히 ‘호풍미’와 ‘진율미’에서 관찰된 항산화 활성의 증가는 저온 스트레스에 대한 내성이 더 강한 품종에서 나타나는 특성으로 해석될 수 있다. 반면, ‘소담미’에서 DPPH 라디칼 소거 활성의 변화가 없었다는 것은 해당 품종이 저온 스트레스에 대한 항산화 방어 능력이 상대적으로 약하다는 것을 나타낸다. 이는 ‘소담미’가 저온 환경에서 ROS 축적에 의해 더 큰 산화적 손상을 받을 가능성이 있음을 시사하며, 이로 인해 저온에 대한 내성이 낮을 수 있다. 따라서, DPPH 라디칼 소거 활성은 저온 스트레스 하에서 품종 간 내성 차이를 평가하는 중요한 생리적 지표로 활용될 수 있다(Lin et al., 2007).
폴리페놀과 플라보노이드
폴리페놀과 플라보노이드는 식물이 스트레스 조건, 특히 저온 스트레스 하에서 방어 메커니즘을 활성화하는 중요한 2차 대사물질로 알려져 있다(Di Ferdinando et al., 2012; Yadav, 2010). 이들 항산화 물질은 활성산소종(ROS)을 제거하여 산화적 손상을 방지하고, 세포를 보호하는 역할을 한다. 본 연구에서 세 가지 품종(‘소담미’, ‘호풍미’, ‘진율미’)을 대상으로 한 실험 결과, 낮은 온도에서 생장한 식물체에서 폴리페놀과 플라보노이드 함량은 품종별로 차이를 보였다.
폴리페놀함량의 경우, 21/17°C 처리에 비하여 저온 처리(19/15℃, 17/13℃) 시 호풍미’와 ‘진율미’의 폴리페놀 함량은 각각 47%에서 90%, 42%에서 47% 증가한 반면, ‘소담미’는 6%에서 15% 감소하였다(Fig. 7). 이러한 결과는 저온 스트레스 하에서 폴리페놀의 합성이 저온 저항성에 중요한 역할을 한다는 점을 시사한다. ‘호풍미’와 ‘진율미’는 폴리페놀 함량의 증가를 통해 저온에 따른 산화 스트레스를 완화할 수 있었던 반면, ‘소담미’는 폴리페놀 함량이 감소하여 저온에 대한 내성이 약해진 것으로 보인다. 폴리페놀은 항산화제로 작용하여 ROS를 제거하고 세포 손상을 방지하는데 중요한 역할을 하므로, 폴리페놀 함량의 증가는 저온에 대한 내성 증가와 밀접한 관련이 있다(Lin et al., 2007).
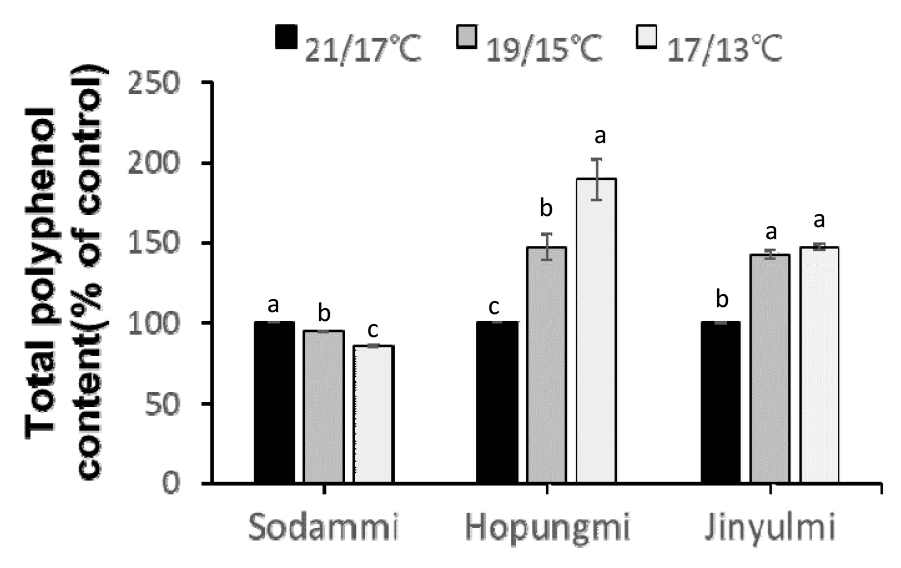
Fig. 7.
Total polyphenol content in the shoots of tthree sweetpotato cultivars under different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) for 35 days. Values (mean ± SE) with different letters within the same cultivar indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test. The control refers to the sample cultivated at 21/17°C.
플라보노이드 함량 역시 저온 스트레스에 따라 유사한 경향을 보였다. ‘호풍미’와 ‘진율미’에서 플라보노이드 함량이 저온 처리에 따라 증가한 반면, ‘소담미’는 감소하는 경향을 보였다(Fig. 8). 플라보노이드는 폴리페놀과 마찬가지로 항산화 특성을 가지며, 저온 스트레스에 대응하기 위한 주요 방어 물질로 작용한다(Di Ferdinando et al., 2012; Yadav, 2010). ‘호풍미’와 ‘진율미’의 플라보노이드 함량 증가는 이들 품종이 저온에 강한 적응성을 가지고 있음을 나타내며, 이는 저온에서 식물의 생존과 생장에 긍정적인 영향을 미친다.
본 연구를 통해 ‘호풍미’와 ‘진율미’는 생육 초기의 저온 스트레스 조건에서도 상대적으로 높은 엽록소 함량과 항산화 활성, 폴리페놀 및 플라보노이드 함량을 유지하여 저온에 대한 내성을 갖는 반면, ‘소담미’는 이러한 특성에서 감소를 보여 저온에 민감한 반응을 보이는 것으로 나타났다. 이는 남부지역에서 4월 정식 시 저온에 대한 대응 품종 선발에 유용한 자료가 될 것으로 판단된다.
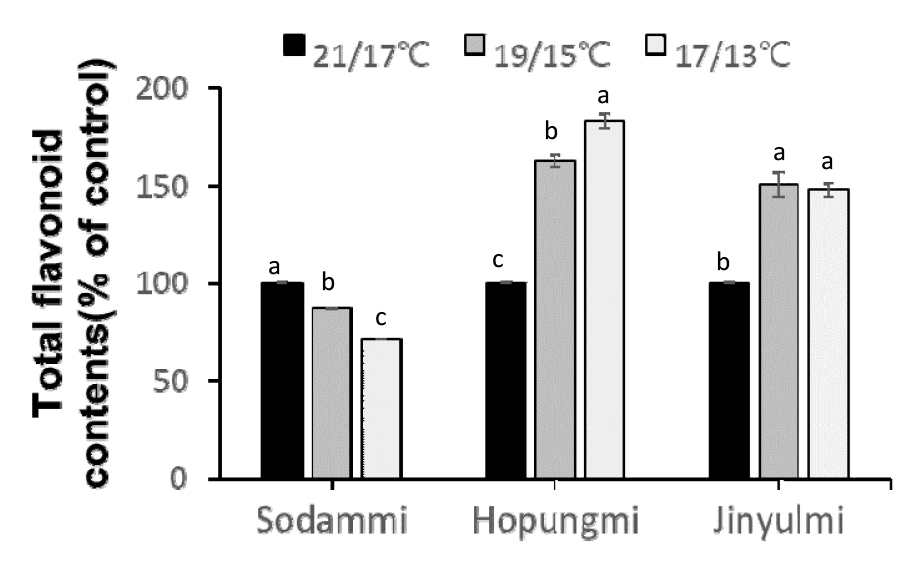
Fig. 8.
Total flavonoid content in the shoots of tthree sweetpotato cultivars under different temperature treatments of 21/17℃, 19/15℃, and 17/13℃ (14 h day temp. /10 h night temp.) for 35 days. Values (mean ± SE) with different letters within the same cultivar indicate significant differences at P < 0.05, as determined using Duncan’s multiple range test. The control refers to the sample cultivated at 21/17°C.
적 요
고구마는 열대성 작물로서 저온에 매우 민감하다. 특히 생육 초기의 저온은 뿌리 수 감소, 생장 지연 등으로 괴근 형성에 영향을 주어 최종 수량을 결정하는 데 영향을 미친다. 신품종 ‘호풍미’, ‘소담미’와 조기 재배용으로 재배 되고 있는 ‘진율미’ 품종을 이용하여 정식 후 생육 온도 차이에 따른 생육 및 2차 대사물질의 변화 양상에 대한 기초자료를 얻고자 생장상에서 수행하였다.
1.저온에서는 세 품종 모두에서 생장 지표에 부정적인 영향을 미쳤으나, 그 정도는 품종에 따라 차이가 있었다. ‘소담미’는 저온에서 상대적으로 생장이 불량 하였으며, 이는 저온 스트레스에 민감함을 나타낸다. 반면, ‘호풍미’와 ‘진율미’는 저온에서도 비교적 양호한 생장 특성을 보여 ‘소담미’에 비해 저온 스트레스에 대한 내성이 더 강할 수 있음을 시사하며, 남부지역에서 온도가 낮은 4월 정식 재배에 유리할 것으로 판단된다.
2.처리 온도가 낮을수록 엽록소 함량이 감소하였으며, ‘소담미’는 엽록소 함량 감소 폭이 가장 컸으며,‘호풍미’와 ‘진율미’는 상대적으로 엽록소 함량이 유지되어 저온에서도 광합성 능력을 어느 정도 유지할 수 있었다.
3.‘호풍미’와 ‘진율미’는 저온에서 DPPH 라디칼 소거 활성이 증가한 반면, ‘소담미’는 유의미한 변화가 없었으며, 폴리페놀 및 플라보노이드 함량 역시 ‘호풍미’와 ‘진율미’는 증가한 반면, ‘소담미’는 감소한 것으로 보아, ‘호풍미’와 ‘진율미’는 저온 스트레스에 대한 항산화 방어 능력이 강한 것으로 생각된다.
