

서 론
재료 및 방법
시험 재료 및 재배 조건
농업형질 조사
밀가루 품질 분석
등숙시기별 유전자 발현 양상 분석
통계 분석
결과 및 고찰
생육기간 온도구배터널의 온도 변화
온도상승에 따른 빵용 밀 수량 및 품질 특성 변화
온도상승에 따른 단백질과 전분 생합성 관련 유전자 발현 변화
적 요
서 론
밀(Triticum aestivum L.)은 전세계적으로 약 20%의 단백질과 약 70%의 칼로리를 제공하는 가장 중요한 작물로, 온대 지역, 온난 다습환경에서부터 한랭 건조한 지역에까지 광범위한 환경 적응성을 지닌 작물이다(Acevedo et al., 2002; Li, 2013; FAO, 2018; Khan et al., 2020). 그러나 밀은 열 스트레스에 매우 민감하기 때문에 기상이변으로 인한 온도 증가로 세계의 많은 지역에서 밀 생산성을 감소가 보고되었다(Hansen et al., 2012; Gupta et al., 2013; Fontana et al., 2015; Mueller et al., 2015). 밀 파종 후 출현 최저온도는 0°C, 분얼 적정온도는 13~18°C, 출수 후 적정온도는 14~26°C로 알려져 있으며, 우리나라를 포함한 아시아 지역의 등숙기 평균기온이 3~4°C 상승 시 수량은 10~50% 감소하는 것으로 예상하고 있다(Nuttallet al., 2018; Khan et al., 2020). 등숙기의 고온 스트레스는 등숙속도를 가속화하여 수량 및 천립중 감소를 일으키는데, 국내에서도 평균 온도 상승 시 금강밀의 출수기가 빨라졌으며, 수수와 천립중 감소로 수량이 감소하였다(Dwivedi et al., 2017; Bergkamp et al., 2018; Jeong et al., 2018; Mahdavi et al., 2021).
밀 생육기간 온도 상승에 관련된 다양한 생리적인 연구가 수행되었는데, 온도구배터널(Temperature Gradient Tunnels, TGT)는 포장 상태와 비슷한 환경하에서 온도 상승 조건을 형성할 수 있는 장점이 있다(Hadley et al., 1995; Horie et al., 1995). TGT를 이용한 평균 온도상승으로 생육 및 등숙기간이 단축되면서 건물중과 면적당 종실수 감소로 수량 감소가 발생하였으며, 생육기간 중 개화 전 최고 온도가 31°C 이상인 경우에는 종실수 감소로 인한 수량 감소가 크게 발생하였다(Fischer & Maurer, 1976; Batts et al., 1996; Wheeler et al., 1996). 등숙기 고온 처리로 성숙기 단축이 발생하여 종실 최대 건물중이 감소하였고, 전분 축적이 적온에 비해 빠르게 종료되었다(Altenbach et al., 2003).
온도상승은 품질에도 영향을 미치는데, 고온은 전분합성을 저해하고 전분 과립 형태도 변화시키며, 아밀로스와 아밀로펙틴 비율을 변화시킨다(Liu et al., 2011; Dwivedi et al., 2017; Li et al., 2018; Kumari et al., 2020). 고온에 의한 단백질 함량 증가는 침전가와 반죽 시간을 증가시킬 뿐만 아니라 고온에 의해서 글리아딘/글루테닌 비율 변화로 글루텐의 특성이 변하게 된다(Tahir et al., 2006; Tao et al., 2018). 이러한 변화는 고온 스트레스에 관련된 다양한 유전자 발현 조절이 관련된 것으로 알려져 있지만(Kino et al., 2020; Kumari et al., 2020; Rangan et al., 2020), 온도상승에 따른 종실 등숙시기별 유전자의 발현 변화에 대한 연구는 미흡한 실정이다.
TGT을 이용한 온도상승에 따른 밀의 생리 반응 연구가 일부 수행되었으나, 우리나라와는 다른 재배 환경에서 농업적 생육 특성에만 국한되었기 때문에 국산밀을 이용한 생육 특성 및 품질 변화 연구가 필요하다. 본 연구에서는 기후변화에 따른 국산밀의 생육기간 온도상승에 대응하기 위해 온도 상승에 따른 국산 빵용 밀의 수량과 품질 변화와 관련 유전자 발현을 평가하여, 온도 상승이 밀 생육에 미치는 영향을 검토하고 향후 기후변화 대응을 위한 기초 자료로 활용하고자 수행하였다.
재료 및 방법
시험 재료 및 재배 조건
본 연구는 빵용 밀 품종인 백강과 조경을 이용하였으며, 2020년 11월 10일에 온도구배터널(TGT; Temperature Gradient Tunnels, 25 m × 2.4 m) 4개동에 각각 14 kg/10a, 휴폭 25 cm 간격으로 4줄씩 줄뿌림 하였다. 시비량은 N-P2O5-K2O를 표준시비량인 10a당 9.1-7.4-3.9 kg 사용하였으며, 관수는 포장 수분 상태를 고려하여 점적관수를 통해 관수하였다. TGT 입구는 공기가 안으로 유입될 수 있도록 개방되어 있으며, 가장 안쪽에는 두 개의 환풍기로 공기를 순환시킬 수 있는 구조로 되어있어 안쪽으로 들어갈수록 측정 지점 온도가 대략 1°C씩 증가하는 시스템이다. TGT의 온도 측정은 입구로부터 약 3.4 m (T0), 9.4 m (T1), 15.4 m (T2), 21.4 m (T3) 지점에 온도센서(SP-110, Apogee instruments, USA)를 설치하여 대조(T0)와 +1°C (T1), +2°C (T2), +3°C (T3) 지점의 온도변화를 기록하였다.
농업형질 조사
농업 형질 조사는 농촌진흥청 농업 과학 기술 조사 분석 기준(RDA, 2012)에 준하여 실시하였다. 출수기는 전체 단위 면적당 이삭 수 중에 70%가 출수한 날로 하였으며, 일수립수는 생육이 균일한 장소에서 무작위로 20수를 채취하여 탈곡 후 조사하였다. 수량은 수확 탈곡 건조 후(수분 14% 기준) 설립을 제외하고 종실을 평량하여 10a당으로 환산하여 조사하였다.
밀가루 품질 분석
밀가루 품질 분석을 위해 각 조사구에서 수확한 원맥 1 kg을 이용하여 수분 14%로 가수처리한 후 제분기(Buhler MLU-202, Bühler, Uzwil, Switzerland)를 이용하여 제분하였다. 회분, 단백질 함량(%)과 침전가는 AACCI Approved Methods 08-01.01, 46-30.01, 56-61.02 (AACCI, 2010)에 준하여 측정하였다. 밀가루의 아밀로스 함량과 총 전분 함량은 각각 Williams et al. (1970)의 방법과 Total Starch Assay Kit (K-TSTA, Megazyme, Chicago, IL, USA)를 이용하여 분석하였다.
등숙시기별 유전자 발현 양상 분석
생육기간 중 온도상승에 따른 글리아딘, 글루테닌, 전분 합성 및 대사 관련 유전자 발현을 등숙기 종자 발달 시기에 따라 분석하였다. 등숙시기별 종자는 T0와 T3 지점에서 출수 후 15일부터 50일까지 5일 간격으로 수수의 중간부분에서 채취하였다. 이후 액체질소를 이용하여 종자를 곱게 분쇄하였고, RNeasy Plant Mini Kit (Qiagen, Hilden, NRW, DE)를 이용하여 RNA를 추출하였다. RNA는 NanoDrop™ 2000/2000c Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA)를 이용하여 정량하였고, RNA to cDNA EcoDry™ Premix (Oligo dT) Kit (Takara, Moriyama, JP)를 사용하여 cDNA를 합성하였다. 유전자 발현 분석에 사용한 프라이머 정보는 Table 1과 같다. 유전자 발현 분석은 TB Green® Premix Ex Taq™ II (Tli RNaseH Plus) (Takara, Moriyama, JP)를 사용하여 Real-Time PCR System (Bio-Rad, CF, CA, USA)를 이용하여 수행하였다.
Table 1.
Oligo nucleotide sequences for qRT-PCR analysis in developing grains.
통계 분석
모든 시험은 3회 반복 수행하여 결과를 획득하였으며, 통계 분석은 SPSS 23 (SPSS Inc., Illinois, USA) 통계 패키지를 이용하였다. 통계적 검증은 일원분산분석(ANOVA)을 수행하였고, Duncan 다중범위검정을 실시하여 p<0.05 수준에서 유의성을 검정 하였다.
결과 및 고찰
생육기간 온도구배터널의 온도 변화
밀 생육기간 중 TGT 각 지점(T0~T3)의 월평균 온도 차이는 Table 2와 같다. TGT 내부의 월평균 온도 차이는 각 지점에 따라 약간의 차이는 보이나 터널 입구(T0)를 기준으로 각 지점에서 대략 +1°C (T1), +2°C (T2) 그리고 +3°C (T3)로 온도구배가 잘 형성되었음을 알 수 있었다. 일평균 온도 역시 TGT의 각 지점에서 온도구배가 잘 형성되었으며, T0 지점은 노지 온도 대비 최저 0.2°C에서 최대 1°C 정도 높게 유지되어(Fig. 1) 실제 재배 온도 조건과 큰 차이는 보이지 않았다.
Table 2.
Mean temperature and thermal difference of the region in the temperature gradient tunnels (TGT). Thermal difference indicates the difference between the elevated temperature (T1, T2, T3) and T0.
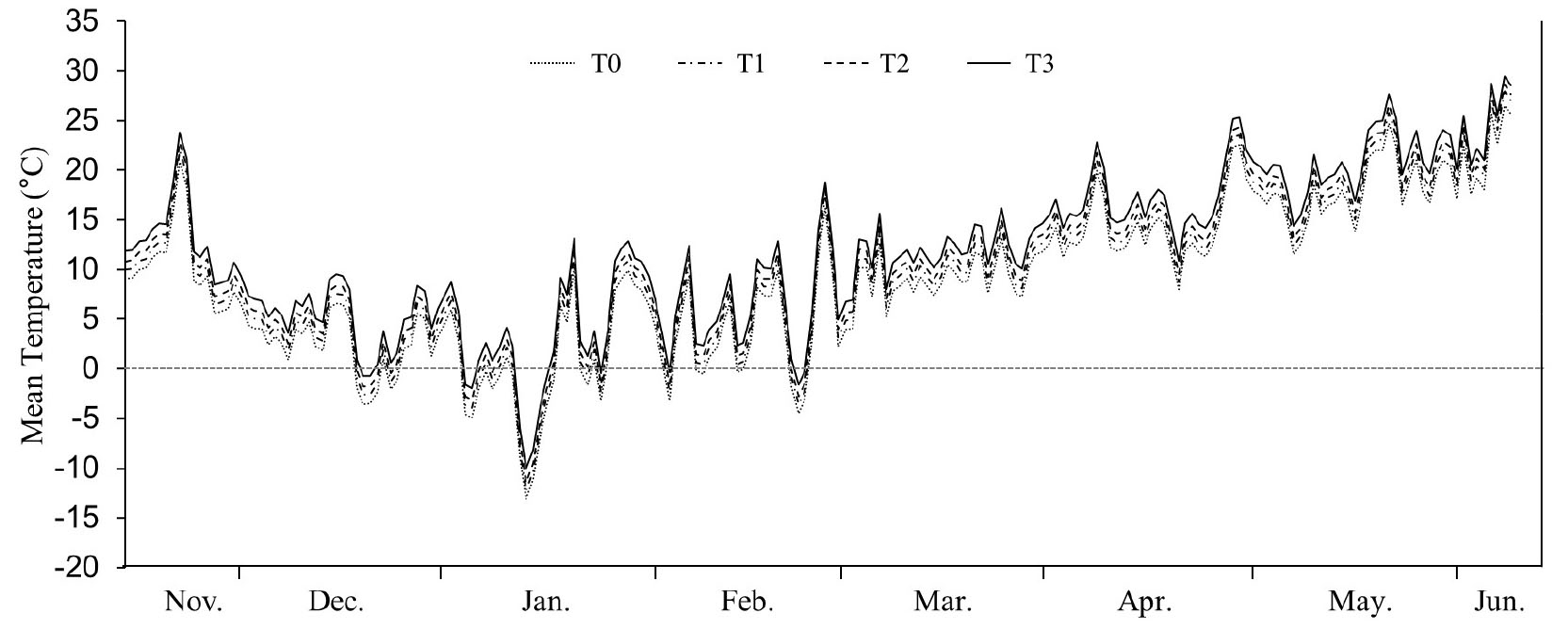
온도상승에 따른 빵용 밀 수량 및 품질 특성 변화
작물의 생육 기간은 온도와 밀접한 관련이 있다. 온도상승은 발육을 촉진하여 생육기간을 단축시키며 작물의 발달에 변화를 초래하여 생산성에 영향을 미친다. 본 연구에서는 TGT을 이용하여 온도상승에 따른 빵용 밀의 생육 특성을 조사하였다(Table 3). 백강과 조경 두 품종 모두 연차 간 차이는 보이나 밀 생육기간 중 온도상승에 따라 출수기가 단축되었으며, 1°C 상승함에 따라 출수기는 대략 3일 정도 단축되었는데, TGT의 온도 상승에 따른 출수기 단축은 기존 결과와 같았다(Fischer & Maurer, 1976; Batts et al., 1996; Wheeler et al., 1996).
Table 3.
Growth characteristics of wheat in response to elevated temperature during their growing season.
온도상승에 따른 일수립수는 백강과 조경 품종 간 차이는 보이지 않았으나 온도가 상승함에 따라 감소하였다. 특히 T3 조건에서 일수립수는 백강이 25.8개, 조경은 25.7개로 T0 (백강-30개/조경-30.5개) 대비 14.0%와 15.7%가 감소하였다. 종실 수량은 T0 조건에서 백강이 558 kg/10a, 조경이 551 kg/10a로 두 품종 간 큰 차이는 보이지 않았으며, 온도상승에 따라 두 품종 모두 감소하였다. 백강과 조경 두 품종 모두 T1 조건에서는 수량 감소를 보이지 않았으며, T2 조건에서 각각 19.7%와 16.9%, T3 조건에서 25.6%와 15.8%의 감소율을 보여 백강이 조경보다 수량 측면에서 온도상승에 민감함을 알 수 있었다. 등숙기 중 온도상승은 종실의 등숙을 가속화하여 천립중과 수량을 감소시킨다고 알려져 있으나, 온도상승 정도에 따라 종실 발달과 수량에 미치는 영향은 달라진다(Khan et al., 2020). 낮은 수준의 온도상승은 등숙속도와 종실 발달을 개선하지만 높은 수준의 온도상승은 등숙률과 종실 발달을 저해한다고 알려져 있다(Sofield et al., 1977; Dias & Lidon, 2009). 본 연구에서는 생육기간 온도상승이 백강과 조경 두 품종 모두 일수립수의 감소로 인해 종실 수량이 결정된 것으로 판단되며, 이는 등숙기와 같은 특정 시기에서의 온도 처리나 온도상승의 정도에 따라 기존 결과와는 영향이 다르게 나타난 것으로 판단된다.
온도상승은 밀의 생육 및 수량 뿐만 아니라 전분과 단백질 함량과 조성 등 품질 특성에도 영향을 미친다(Spiertz et al., 2006). 밀가루에서 단백질은 가공 적성과 용도에 영향을 미치며, 전분은 가공 식품의 식미에 영향을 미치는 주요한 요인으로, 단백질의 함량과 글루텐 단백질의 질적 특성 그리고 전분의 함량과 아밀로스 결정 구조 등에 의해 가공 식품의 특성이 결정된다(Baik, 2010; Sharma et al., 2020). 온도상승에 따른 빵용 밀의 품질 특성 변화는 Table 4와 같다. 회분은 배유부에는 적고 외피 부분에 많이 존재하기 때문에 제분율에 따라 밀가루의 회분 함량이 달라지며, 일반적으로 회분 함량이 낮을수록 밀가루의 품질이 높게 평가된다(Hinton, 1959). 백강과 조경의 회분 함량은 T0 조건에서 0.43%과 0.42%로 나타났으며, 두 품종 모두 온도상승에 따른 회분 함량의 변화는 보이지 않았다.
Table 4.
Quality characteristics of wheat in response to elevated temperature during their growing season.
온도상승에 따른 단백질 함량은 T0 조건에서 백강과 조경이 11.8%와 11.6%로 나타났으나, 온도가 상승함에 따라 증가하여 T3 조건에서는 백강이 15.0%, 조경은 15.4%로 나타났다. 글루텐 단백질의 함량과 강도를 간접적으로 알아볼 수 있는 침전가는 온도상승에 따라 증가하여 단백질 함량 증가와 일치하는 경향을 보였다. 전분의 아밀로스와 아밀로펙틴 함량은 국수의 식미에 영향을 미치는 주요 요인으로, 백강과 조경 두 품종의 총 전분 함량은 T3 조건에서 유의하게 감소하여, 전분 구성성분의 함량 변화에 대한 추가적인 연구가 필요할 것으로 생각된다. 온도상승에 따라 단백질 함량은 증가한 반면 총 전분 함량은 감소하였는데, 이러한 결과는 온도상승에 따른 전분 합성은 단백질 합성에 비해 온도변화에 민감하며(Spiertz et al., 2006), 단백질 함량 감소 대비 전분 함량 감소가 많았기 때문일 것이다.
밀 생육기간 중 평균 온도상승은 생육 기간과 등숙기간을 단축시키는 반면, 수량 감소를 유발한다(Fischer & Maurer, 1976; Batts et al., 1996; Jeong et al., 2018). 또한 고온 스트레스는 엽록소 감소로 인한 광합성 저하와 잎의 노화와 더불어 종실의 등숙기간 감소를 유발한다(Farooq et al., 2011). 백강과 조경 두 품종의 등숙시기별 온도상승에 따른 립중 변화를 분석하기 위해 출수 후 15일부터 5일 간격으로 이삭을 채취하여 등숙시기별 립중을 분석하였다. 백강과 조경 두 품종 모두 T0 대비 T3 조건에서 등숙속도가 빨랐으며, 등숙 완료 시기도 대략 5일가량 단축되었다(Fig. 2). 따라서 빵용 밀 생육기간 중 온도상승은 등숙을 촉진하고 등숙 기간을 단축시키나 품종 간 온도에 대한 생리적 반응에서는 차이가 없는 것으로 판단된다.
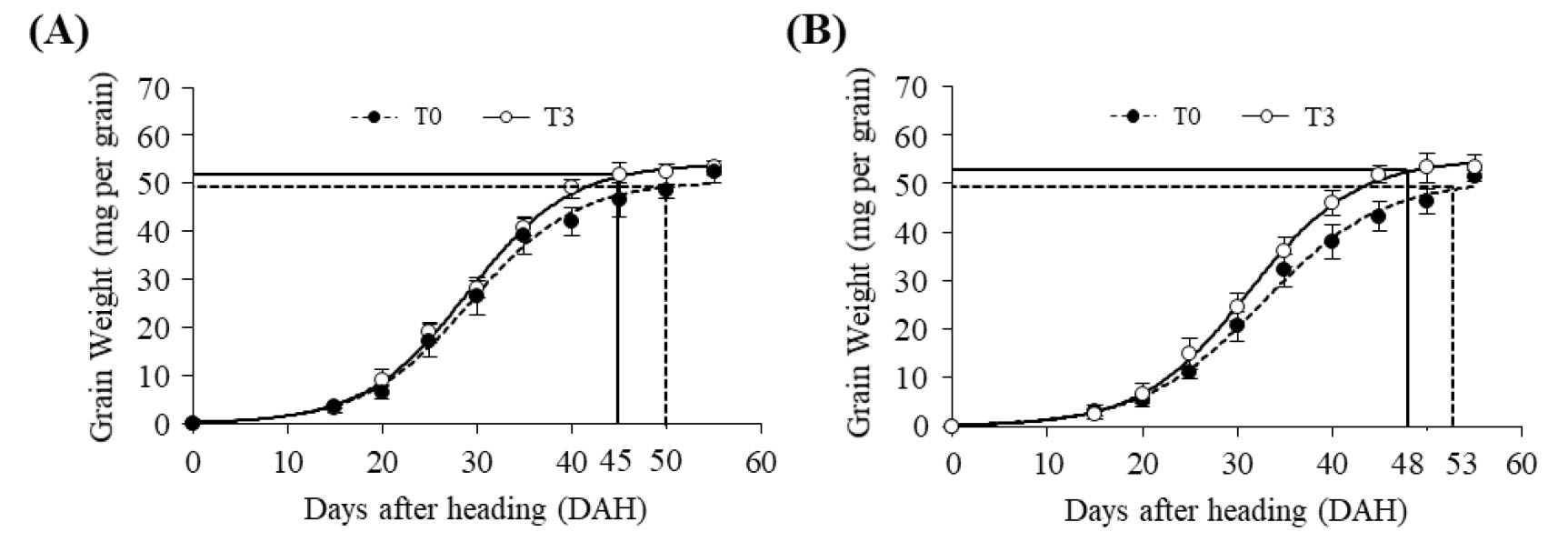
Fig. 2.
Effect of elevated temperature on the grain dry weight of Baekkang (A) and Jokyoung (B) during grain filling period. Solid and dotted lines in the graphs indicate the days from DAH to the maturity of grains. Error bars indicate standard deviation determined from three independent biological replicates.
온도상승에 따른 단백질과 전분 생합성 관련 유전자 발현 변화
밀 생육기간 중 고온 스트레스는 다양한 경로와 관련된 유전자의 발현을 조절한다. 개화 후 고온은 과피 세포벽 신장과 관련된 유전자의 발현을 저해함으로써 종자 무게의 감소를 유발한다(Kino et al., 2020). 등숙 초기와 후기에 고온 민감성 품종 대비 고온 저항성 품종에서 발현이 증가되는 유전자군이 확인되었고(Rangan et al., 2020), genome-wide 연구를 통해 고온과 건조 스트레스 시 발현에 변화를 보이는 열 충격 단백질(HSP, heat shock protein) 관련 유전자와 전사 인자 관련 유전자들이 확인되었다(Kumari et al., 2020). 또한 고온 스트레스는 종자 발달 단계에서 전분 생합성 경로, 전분 이동, 전사 인자 그리고 저장 단백질 관련 유전자 발현 등을 다중 경로를 통해 조절한다고 알려져 있다(Chunduri et al., 2021). 글리아딘을 암호화하는 유전자 중 γ-와 ω-type 유전자는 Gli-1 (Metakovsky et al., 2006), α-type 유전자는 Gli-2 유전자좌(Masci et al., 2002)에 존재한다. 글루테닌 합성에 관여하는 유전자는 Glu-D1Dx2와 Glu-D1Dx12가 있으며, 글루텐 단백질 응집에 관여하는 유전자는 PDIL2-1, PPIase 및 SUMO1, 종자 발달 동안 전사 인자로 기능하는 PBF, SPA 및 GAMyb 유전자가 있다(Song et al., 2020). 전분은 포도당-1-인산으로부터 5가지 유형의 대사 과정 유전자의 조절을 통해 합성된다: ADP-glucose pyrophosphorylase (AGPase), starch synthase (granule-bound starch synthase, GBSS; soluble starch synthase, SS), starch-branching enzyme (SBE), starch-debranching enzymes (DBEs) (isoamylase, ISA; pullulanase, PUL), 그리고 starch-degrading enzymes (starch phosphorylase, Pho; starch-disproportionating enzyme, DPE; α- and β-amylases) (Singh et al., 2015).
종실 등숙시기별 온도상승에 따른 저장단백질과 전분 합성 및 대사 관련 유전자의 발현을 분석하기 위해 real-time RT-PCR을 수행하였다(Fig. 3). 글리아딘 생합성 관련 유전자인 α-Gli (α-type), g1Gli (γ-type)과 Q18, Q19 (ω-type)의 발현 분석 결과, 백강과 조경 두 품종에서 발현양과 발현 양상이 약간의 차이는 보이지만, T0 조건에서는 출수 후 15일부터 40일까지 발현이 증가하다 출수 후 45일에는 발현이 거의 되지 않았다. 반면 T3 조건에서는 출수 후 45일에도 발현이 높게 유지됨을 확인하였다(Fig. 3A). 다음으로 종자 발달과정 중 전사 인자로 글루테닌 대사에 관여하는 유전자인 SPA와 PBF, 고분자 글루테닌 생합성에 관여하는 유전자 GluDx와 GluDy의 온도상승에 따른 발현 변화를 분석하였다. 백강과 조경 두 품종에서 발현 정도와 양상에 차이는 보이나 T0 조건에서 출수 후 15일부터 40일까지 발현이 증가하였고 출수 후 45일에는 발현이 거의 되지 않았다. 반면 T3 조건에서는 T0 조건에 비해 발현양은 낮지만 출수 후 45일까지 발현이 유지되어(Fig. 3B) 글리아딘 합성 관련 유전자와 유사한 발현 양상을 보였다. 전분 합성 관련 유전자인 GBSSI, ISA-1, SSI와 SBEII는 백강과 조경 두 품종에서 발현 양상의 차이는 보이나 등숙기 동안 T3 조건과 비교하여 T0 조건에서 전반적으로 높은 발현양을 보여(Fig. 3C) 글리아딘과 글루테닌 생합성 및 대사 관련 유전자 발현과는 다른 경향을 보였다. 온도상승 시 백강과 조경 두 품종에서 단백질 함량이 증가하고 총 전분 함량은 감소하였는데(Table 3), 이러한 결과는 온도상승 시 단백질 합성은 증가하고 전분 합성은 감소한다는 기존의 연구결과와 일치한다(Spiertz et al., 2006). 이러한 결과들을 토대로 온도상승에 따라 글리아딘과 글루테닌 합성 및 대사 관련 유전자의 발현 증가로 인해 단백질 함량이 증가하고, 전분 합성 관련 유전자의 발현 감소로 인해 총 전분 함량이 감소했을 것으로 판단된다.
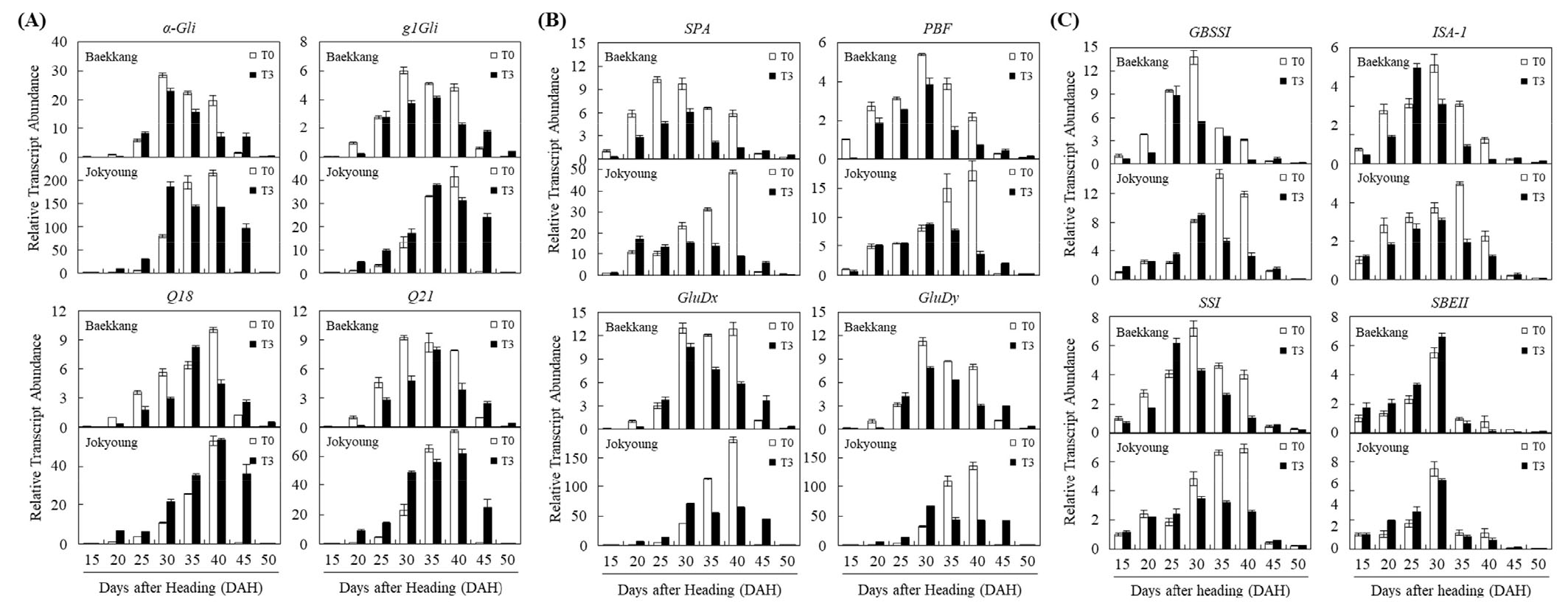
Fig. 3.
Expression profiles of gliadin synthesis-related genes (A), glutenin metabolism-related genes (B) and starch synthesis-related genes (C) grown under the control (T0) and elevated temperature (T3) during grain filling period. Error bars indicate ±SD determined from three independent biological replicates. α-Gli, Encoding α-type gliadin gene; g1Gli, Encoding γ-type gliadin gene; Q18 and Q21, Encoding ω-type gliadin gene; SPA, Storage protein activator; PBF, Prolamin-box binding factor; GluDx, High molecular weight glutenin subunit; GluDy, High molecular weight glutenin subunit; GBSSI, Granule-bound starch synthase I; ISA-1, Isoamylase; SSI, Soluble starch synthase I; SBEII, Starch-branching enzyme II.
적 요
본 연구에서는 온도상승에 따른 국내산 빵용 밀의 수량과 품질 변화 연구를 위해 TGT을 이용하여 생육기간 중 정상 온도 대비 1-3°C 증가한 조건에서 수량구성요소와 수량 그리고 밀가루 품질 및 분자생리적 요인을 분석함으로써 밀 생육기 전반에 미치는 온도상승의 영향을 연구하였고, 연구 결과는 다음과 같다.
1. 밀 생육기간 중 평균온도가 1°C 상승함에 따라 백강과 조경 두 품종 모두 출수기는 3일 정도 단축되었고, 일수립수가 감소하였으며, 온도가 상승함에 따라 수량이 감소하였다.
2. 밀가루 품질 분석 결과 아밀로스와 회분 함량은 백강과 조경 두 품종 모두 온도상승에 의한 영향을 받지 않았으나, T0 조건 대비 T3 조건에서 단백질 함량은 증가하였고 총 전분 함량은 감소하였으며,
3. 등숙기 중 글리아딘과 글루테닌 합성 및 대사 관련 유전자 발현 분석 결과 백강과 조경 두 품종 모두 종자 발달 후기에서 T0 대비 T3 조건에서 발현양이 높았으며, 전분 합성 관련 유전자 발현양은 빠르게 감소하였다.
4. TGT을 이용한 국내산 빵용 밀의 생육기 전반에 미치는 온도상승이 수량과 품질에 영향을 미쳤으며, 관련 유전자 발현 분석은 향후 기후변화 대응을 위한 기초 자료로 활용할 수 있을 것이다.
