

식물유전자원을 안전하게 보존하기 위하여 건조된 종자 를 저온 저장고에 보존하지만 생명체는 시간의 경과에 따라 퇴화가 진행되며 이는 불가역적 생리현상(Delouche & Baskin, 1973)이다. 따라서 각 국의 종자은행에서는 보존 중인 자원 의 발아력 저하 시 재증식을 통해 활력이 높은 자원으로 갱 신코자 지속적인 활력모니터링을 실시하고 있다. 대부분의 종자은행에서는 활력모니터링을 저장조건에 따라 -18~-20°C 저장고 보존 자원의 경우 10년 주기로, 1~5°C 저장고 보존 종자는 5년 주기로 실시하고 있다(FAO/IPGRI, 1994).
종자의 저장력을 평가하기 위하여 종자를 고온(41°C) 다습 (약 100% RH) 조건으로 3~4일간 처리하는 노화촉진(accelerated ageing; AA)검사법을 제안하였는데(Delouche & Baskin, 1973), 이는 종자세 검사 및 포장에서 유묘의 출현력을 검사하는 방법으로도 활용되고 있다.
Sato (1991)는 일본의 종자은행에 보존 중인 벼 유전자원 의 저장수명을 알아보기 위해 노화촉진처리(AA)를 한 결 과, 논벼가 밭벼에 비해 활력저하 속도가 빠른 것을 확인하 고 종 내에서도 종자수명의 차이가 있으며, 노화촉진처리를 통해 신속하게 종자수명을 예측할 수 있음을 보고한바 있다.
또 하나의 종자 저장력 평가 방법으로 퇴화조절(controlled deterioration; CD)처리가 있다. 이 방법은 노화촉진(AA) 처 리와는 달리 종자의 수분함량을 일정 수준으로 조절한 다음 동일한 종자수분함량을 유지시키며 고온(40~45°C) 조건에 서 퇴화 처리를 하는 것이다(Matthews, 1998).
최근에 와서 Lee et al.(2002, 2006)은 건열저항성이 종자 의 수명과 유의한 상관관계를 가지므로 화곡류의 종자수명 을 측정하는 간접지표로 활용 가능성을 제시한바 있다. 건 열처리는 일반적으로 종자의 휴면타파(Dadlanni & Seshu, 1990), 종자감염 병원균 및 선충 방제(Clear et al., 2002; Nakagawa & Yamaguchi, 1989)를 위한 방법으로 이용되어 왔으며 보통 50~80°C에서 처리한다. Lee et al.(2006)이 제 안한 건열처리(dry-heat treatment; DHT)는 종자수분함량이 약 8%가 되도록 사전건조 후 90°C에 처리하는 것으로 벼를 대상으로 24시간의 건열처리 후 발아율은 실온에서 6~54개 월 보관 후의 발아율과 고도의 유의한 정의 상관관계를 보 였다고 하였다.
또한 인위노화처리의 방법인 노화촉진(AA)처리와 퇴화 조절(CD)처리를 통해 여러 작물의 종자세 검사(Hampton & TeKrony, 1995; Kim et al., 1987a)와 포장 출현율 및 종 자의 저장력 예측(TeKrony et al., 2005; Kim et al., 1994) 이 이루어졌다. 작물별 인위노화처리를 통한 종자세와 종자 저장력 예측 연구를 살펴보면 콩(Kim et al., 1987a), 보리 (Kim et al., 1987b), 양파, 당근, 배추과 작물 등 채소작물 (Powell & Matthews, 1981), 레드클로바(Wang et al., 1994), 수수(Ibrahim et al., 1993; Kim et al., 2013) 및 옥수수 (TeKrony et al., 1989)등에 대한 보고가 있었다.
현재까지 벼 종자수명에 대한 연구의 흐름을 정리해 보 면 초기에는 다른 작물과의 비교 차원에서 종 수준에서 종 자수명을 측정하였다. 이후 벼의 생태형에 따라 수명의 차 이가 있음을 발견하였으며, 휴면성, 배유형, 수확시기 및 재 배환경 등이 종자수명에 미치는 영향을 연구하였다. 이들 연구에서는 종자의 수명을 조기에 비교 및 예측하기 위하여 인위노화처리를 실시하였다. 벼의 인위노화처리 방법으로 는 주로 일정한 종자수분함량을 유지시키며 고온에 보관하 는 퇴화조절(CD)처리를 주로 사용하여왔다.
본 연구는 농촌진흥청 국립농업과학원 농업유전자원센터 의 저온 저장고에 보존되어온 벼 유전자원의 종자수명을 살 펴보고, 이들 자료를 바탕으로 효과적인 종자의 저장수명을 예측할 수 있는 방법을 규명하여 벼 유전자원의 활력모니터 링 주기설정 및 갱신 계획 수립에 필요한 기초자료를 제공 하고자 수행하였다.
재료 및 방법
시험 재료
2007년 농업유전자원센터 증식 포장에서 수확한 생태형, 원산지 및 생육특성이 다양하게 포함된 자포니카형 48, 통 일형 10, 인디카형 25와 자바니카형 23 전체 106자원의 벼 핵심집단(Chung et al., 2004, Chung & Park, 2009)을 실온 에 5개월 동안 두며 자연적으로 휴면이 타파되도록 하여 10°C, 상대습도 35%의 저온 저장실에 보관하며 사용하였다.
벼 유전자원의 종자수명을 예측하기 위한 최적의 인위노 화처리 방법을 규명하기 위해서 노화촉진(AA)처리 시험에 는 생태형별 각 6개 품종을 공시하였으며, 퇴화조절(CD)처 리 시험에는 생태형별 각 2개 품종을, 건열처리(DHT) 시험 에는 106개 품종을 공시하였다.
4°C 저장고에서의 벼 생태형별 저장수명을 알아보기 위 해서는 농촌진흥청 농업유전자원센터에 26.5년간 보존된 벼 유전자원 3,066점의 활력검정 성적을 활용하였다.
종자수명 예측 방법 규명
벼 유전자원의 효과적인 종자수명 예측 방법 규명을 위 해 인위노화처리 방법인 노화촉진(AA)처리, 퇴화조절(CD) 처리 및 건열처리(DHT)를 실시하였다.
노화촉진(AA)처리는 철망트레이법(McDonald & Phaneendranath, 1978)으로 플라스틱 사각박스(11.0×11.0×3.5 cm)에 증류수 80ml를 가하고, 철망 트레이에 종자를 단층으로 고루 펼친 다음 사각박스에 넣고 뚜껑을 닫아 41°C 항온기에 12일간 두면서 3일 간격으로 발아검사를 실시하였다.
퇴화조절(CD)처리는 종자를 1, 3 및 6시간 동안 각각 흡습 을 시킨 후 종자를 알루미늄박 봉투에 담아 밀봉하여 25°C 항온실에 하루 동안 보관하며 수분평형을 이루게 한 다음 종자수분함량을 조사하고, 각 흡습처리된 종자를 45°C 항 온기에 3~4일 간격으로 24일간 퇴화처리 기간을 달리한 후 발아시험을 실시하였다. 종자 흡습은 플라스틱 사각박스(11.0× 11.0×3.5 cm)에 증류수 50 ml를 가하고, 철망 트레이에 종 자를 단층으로 고루 펼친 다음 사각박스에 넣고 뚜껑을 닫아 25°C 항온기에 넣어 실시하였다. 종자수분함량(seed moisture content; SMC) 조사는 ISTA (1999)에서 권장하는 고온항 온건조기법에 따라 실시하였다. 각 자원당 2반복으로 실시 하였으며, 반복 간 수분함량의 차가 0.2%보다 크면 재조사 를 실시하였다.
M1: 수분측정 캔의 무게. M2: 건조 전 총무게. M3: 건조 후 총무게
건열처리(DHT)는 수분함량이 9.9±0.2%인 종자를 종이 봉투에 담아 90°C 건조기에서 각각 12, 24 및 36시간 동안 처리하였다. 처리가 끝난 종자는 실온에서 1일간 방치하였 다가 발아검사를 실시하였다. 발아검사는 ISTA (1999)의 ‘International Rules for Seed Testing’에 따라 between paper (BP) 방식으로 50립씩 4반복으로 25°C 항온실에서 실시하 였다.
종자수명(P50)은 발아율이 50%까지 저하되는데 소요되는 기간으로, 처리에 따른 발아율 변화 추세선을 그리고 그 회 귀방정식 y값이 50이 되는 x값으로 구하였다.
자료 분석
모든 성적의 분석은 Microsoft Excel 2010의 함수마법사 와 SAS 통계 소프트웨어(V 9.1)를 이용하였다. 평균값의 비교는 유의수준 5%로 분산분석을 실시하였다.
결과 및 고찰
인위노화처리 방법에는 많은 작물을 대상으로 노화촉진 처리, 퇴화조절처리가 대표적으로 이용되고 있다. 최근 벼 품종의 건열저항성이 종자수명과 고도의 유의한 상관관계 를 갖는다는 보고(Lee et al., 2006)가 있었다. 벼 유전자원 의 종자수명을 효과적으로 예측할 수 있는 방법을 규명코자 노화촉진처리, 퇴화조절처리 및 건열처리를 실시한바, 다음 과 같은 결과를 얻었다.
노화촉진처리
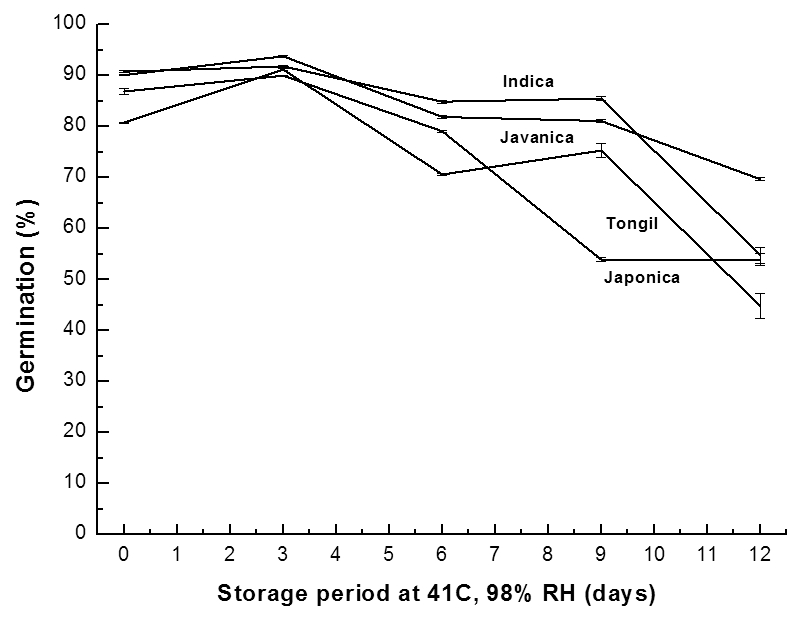
벼 생태형별 각 6품종을 41°C, 98% RH 조건의 노화촉진 처리 기간에 따른 벼의 활력변화는 Fig. 1과 같다. 노화촉진 처리에 따른 벼 종자의 활력저하 경향이 생태형별로 균일하 지 않았다. 그 이유는 고온 과습한 조건에서 곰팡이 등 유해 미생물들의 작용이 큰 영향을 미친 것으로 사료된다. 따라 서 추후 벼 종자의 저장력 평가에 알맞은 처리온도와 처리 시간의 검토가 더 이루어져야 할 것으로 판단된다.
퇴화조절처리
퇴화조절처리를 위한 종자의 수분함량을 조절하기 위해 각 생태형별로 2품종을 공시하여 흡습시간에 따른 종자의 수분함량 변화를 조사하였다(Table 1). 흡습처리 전 생태형 별 수분함량은 인디카형, 자바니카형, 통일형은 각각 9.7~ 9.8%인데 비해 자포니카형 벼는 10.3%로 같은 환경조건에 서 1개월 이상 보존한 것을 고려해 보면 평형수분함량이 상 대적으로 높음을 알 수 있었다. 6시간 흡습한 종자의 생태 형별 수분함량은 자포니카형이 14.8%인데 반해 인디카형 은 15.0%로 초기 수분흡습은 인디카형 벼가 빠르다는 것을 알 수 있었다. 12시간 흡습 후에는 자포니카형과 인디카형 벼의 종자수분함량이 17.8%로 동일하였으며, 48시간 동안 수분을 흡습했을 때는 자포니카형이 25.0%로 인디카형 벼 종자수분함량 23.4%보다 1.6% 높은 수분함량 차이를 보였다.
퇴화조절 처리를 위해 흡습 시간을 1시간, 3시간, 6시간 으로 조절하였을 때 공시 벼 종자의 평균 수분함량은 각각 11.3±0.2%, 12.5±0.3%, 14.4±0.5%였다. 흡습한 종자를 알 루미늄박봉투에 담아 밀봉하여 수분함량변화가 없도록 한 다음 45°C 고온항온기에 두면서 발아율 변화를 살펴본 결 과는 Fig. 2와 같다. 1시간 흡습처리 한 종자의 경우 45°C 고온항온기에 저장한 후 21일째에 생태형간 퇴화정도의 차 이가 뚜렷하게 나타났으며, 3시간, 6시간 흡습 처리구에서 는 각각 14일, 10일째에 뚜렷한 차이를 보였다.
Table 1.
Changes in seed moisture content (%, wb) of rice germplasms after imbibition at 25°C 98% RH.
| Ecotype | Imbibition time (hours) | |||||
|---|---|---|---|---|---|---|
| 0 | 6 | 12 | 18 | 24 | 48 | |
| Indica | 9.7a† | 15.0a | 17.8a | 19.5a | 20.5a | 23.4b |
| Japonica | 10.3a | 14.8ab | 17.8a | 19.8a | 21.2a | 25.0a |
| Javanica | 9.8a | 14.4b | 17.2a | 19.1a | 20.1a | 23.8ab |
| Tongil | 9.8a | 14.5b | 17.4a | 19.3a | 20.9a | 24.9a |
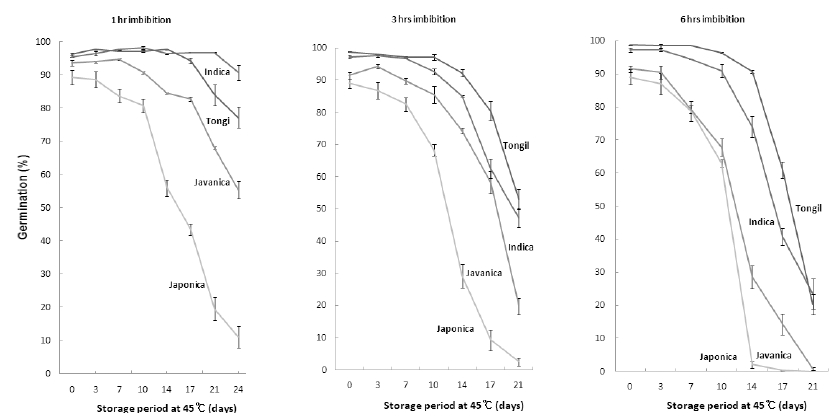
Fig. 2.
Changes in germination percentage after controlled deterioration treatment. The seed moisture contents treated during 1, 3 and 6 hours imbibition were 11.3±0.2%, 12.5±0.3%, 14.4±0.5% respectively. Vertical bars represent standard errors of the means.4°C 저장고에 보존된 자원들의 저장수명을 살펴본 바, 종 자수명은 자포니카형, 자바니카형, 통일형, 인디카형 순으 로 길어졌는데 퇴화조절처리 시에는 1시간 흡습 처리구에 서만 같은 경향을 보이며, 3시간, 6시간 흡습 처리구 에서는 고온항온기 보관 17일까지는 통일형이 인디카형에 비해 퇴 화속도가 늦은 경향을 보였다. 그러나 자포니카형, 인디카 형, 자바니카형 간의 퇴화속도에는 뚜렷한 차이를 보이며 4°C 저장고에 보존된 자원의 생태형별 퇴화 정도와 같은 경 향을 보였다.
흡습시간을 달리하여 45°C 고온항온기에 저장하며 활력 저하를 살펴본 결과, 6시간 흡습 처리구가 1시간 및 3시간 흡습 처리구에 비해 퇴화속도가 빨라 생태형별 자원수를 각 6품종씩으로 늘려 퇴화조절 처리시의 종자수명을 구하였다 (Fig. 3, Table 2).
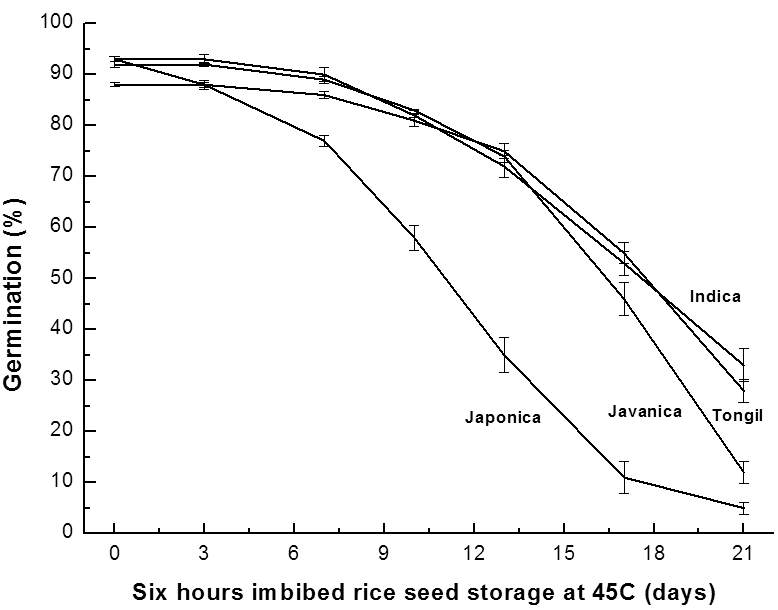
Fig. 3.
Changes in germination percentage of rice ecotypes according to 6 hours imbibed rice seed storage period at 45°C. The seed moisture contents of Indica, Japonica, Javanica and Tongil imbibed during 6 hours were 13.7 ±0.4%, 13.7±0.7%, 13.6±0.5% and 14.0±0.5% respectively. Vertical bars represent standard errors of the means.Table 2.
Estimating seed longevity (P50) and seed germination percentage of rice germplasm accessions through controlled deterioration (6 hours imbibed seed storage at 45°C) using multiple regression analysis.
| Ecotype | Controlled | deterioration | period | (days) | P50 (days) |
|---|---|---|---|---|---|
| 0 | 7 | 13 | 21 | ||
| Indica | 93±4.7a† | 90±8.9a | 72±34.0a | 33±30.6a | 17.9 |
| Japonica | 93±4.0a | 77±14.0b | 35±34.5b | 5±12.9a | 10.8 |
| Javanica | 92±3.8a | 89±5.5a | 74±12.9a | 12±11.8a | 16.4 |
| Tongil | 88±6.0a | 86±4.0ab | 75±9.6a | 28±25.1a | 17.8 |
Fig. 3의 퇴화조절 처리에 따른 발아율 변화를 근거로 추 세선을 그리고 회귀방정식을 구하여 생태형별 평균 종자수 명을 계산하였다(Table 2).
퇴화조절 처리 시 벼 생태형별 활력변이의 회귀방정식을 이용한 종자수명은 자포니카형 벼의 경우 10.8일이며, 인디 카형, 통일형, 자바니카형은 각각 17.9일, 17.8일, 16.4일로 계산되었다.
6시간 흡습한 벼 종자를 45°C 고온 항온기를 이용한 퇴 화조절 처리 시 처리 7일차와 13일차에서는 자포니카형 벼 의 활력저하가 다른 생태형과 유의한 수준으로 차이를 보였 으며, 처리 21일차에는 평균발아율은 5%로 가장 낮았지만 DMRT 분석 시 유의한 차이를 보이지 않았다(p=0.1148). 이는 생태형간 발아율의 차이보다 생태형내의 품종 간 발아 율의 차이가 더 컸기 때문으로 사료된다.
건열처리
벼 종자의 건열처리 후 수분함량 변화는 Table 3과 같다. 건열처리 전 평균 수분함량은 10.3±0.2%이었던 것이 건열 처리 24시간 후에는 생태형별 차이 없이 종자수분함량 2.1% 가 되었다.
Table 3.
Changes in seed moisture content (%, wb) of rice germplasm accessions after dry-heat treatment (90°C, 24 hours).
| Ecotype | No. of accessions | Initial | After dry-heat treatment |
|---|---|---|---|
| Indica | 26 | 10.2 | 2.1±0.1† |
| Japonica | 48 | 10.3 | 2.1±0.2 |
| Javanica | 23 | 10.2 | 2.1±0.2 |
| Tongil | 10 | 10.4 | 2.1±0.2 |
인디카형 및 통일형 벼의 경우 고온(90°C)에서 종자 수분 함량 2.1%까지 건조되어도 발아율이 85% 이상 유지하는 자원이 많은 것으로 보아, 인디카형 벼를 주로 재배하는 열 대지역의 저온 저장시설을 갖추지 못한 곳에서 벼 종자를 오랜 기간 저장하기 위해서는 고온(70°C 수준) 건조를 통해 종자의 수분함량을 낮추어 방습이 가능한 용기에 밀봉 저장 하는 방법을 이용하는 것이 적절할 것으로 사료되었다.
종자수분함량이 10.3±0.2%인 벼 106자원의 종자를 종이 봉투에 담아 90°C 건조기에서 건열처리를 하였을 때 처리 시간에 따른 활력변화는 Fig. 4에서 보는바와 같다. 인디카 형과 통일형 벼는 건열처리 36시간까지도 초기 발아율과 큰 차이를 보이지 않은 반면, 자포니카형 벼는 초기발아율 89%에서 건열처리 36시간 후에는 40%로 낮아졌다. 건열처 리 36시간 후의 활력변화는 4°C 저장고에서 26.5년간 보존 후의 벼 생태형별 퇴화정도와 유사한 경향을 보였다(Na et al, 2014).
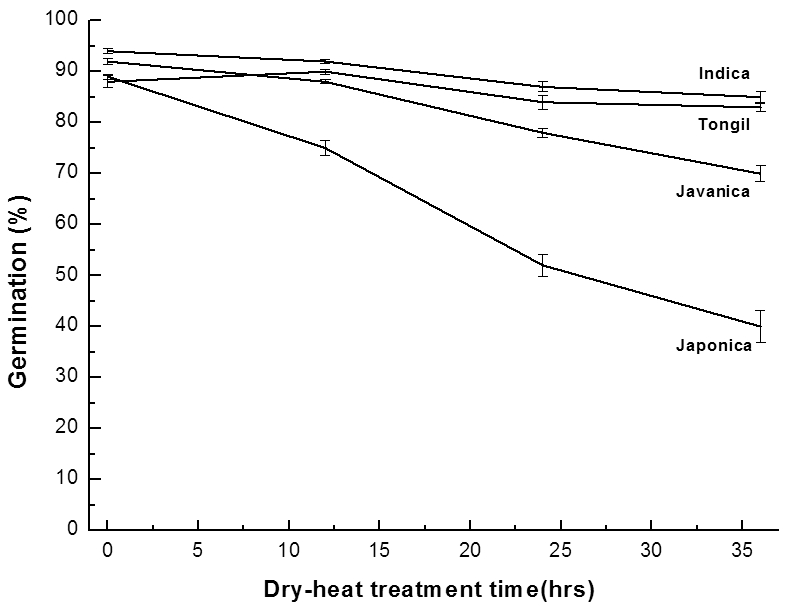
Fig. 4.
Changes of germination percentage of different rice ecotypes after dry-heat treatment (90°C) for various period. Vertical bars represent standard errors of the means.건열처리 경과 시간에 따른 발아율 변화를 근거로 추세 선을 그리고 회귀방정식을 구하여 생태형별 평균 종자수명 을 계산하였다(Table 4). 종자수명은 자포니카형 벼의 경우 27시간이며, 인디카형, 통일형, 자바니카형은 각각 139시 간, 80시간, 57시간으로 계산되었다.
Table 4.
Estimating seed longevity (P50) and seed germination percentage of rice germplasm accessions through dry-heat treatment (90°C) using multiple regression analysis.
| Ecotype | Dry-heat treatment time (hours) | P50 (hours) | |||
|---|---|---|---|---|---|
| 0 | 12 | 24 | 36 | ||
| Indica | 94±3.3a† | 92±5.7a | 87±9.7a | 85±13.9a | 139 |
| Japonica | 89±7.0b | 74±14.2b | 52±19.9b | 40±23.3c | 27 |
| Javanica | 92±5.8ab | 88±8.4a | 78±14.7a | 70±18.0b | 57 |
| Tongil | 88±7.7b | 90±6.9a | 84±13.6a | 83±11.3ab | 80 |
건열처리 36시간 후의 벼 생태형별 활력저하의 정도는 유의한 수준으로 자포니카형이 가장 낮은 발아율을 보였으 며, 자바니카형, 통일형, 인디카형 순으로 발아율 저하가 적 게 나타났는데, 이는 선행된 실험 조사 결과인 4°C 저장고 에 보존 되었던 벼 유전자원의 생태형별 종자수명(Na et al, 2014)과 같은 경향을 나타내는 결과이다.
건열처리를 통한 종자수명 예측
벼 유전자원을 종자은행에 저장하기 전에 각 자원별로 효과적인 종자수명 예측 방법을 규명코자 노화촉진처리, 퇴 화조절 및 건열처리를 실시한바, 건열처리가 가장 간편하면 서도 짧은 시간내에 대량으로 처리할 수 있으며, 저온 저장 고 보존자원의 종자수명과 유사한 경향을 보여 주었다. 특 히 36시간 건열처리 시 벼 생태형별 활력저하의 정도가 유 의한 차이를 보였다. 따라서 벼 유전자원을 종자은행에 저 장하기 전에 종자수명을 예측하기 위한 최적의 방법은 건열 처리(90°C, 36시간)법으로 사료된다.
건열처리가 종자은행에 보존하는 벼 유전자원의 종자수 명 예측 방법으로 적합함을 다음 실험으로 재확인할 수 있 었다. 건열처리 후 최종발아율을 Microsoft의 QUARTILE 함수를 이용하여 사분위수를 구하고 발아율에 따라 4개의 분류군으로 나누었다(Table 5). Ⅰ군은 평균 발아율이 22% 로 아주 낮은 건열저항성을 보였으며, Ⅳ군은 평균 발아율 이 92%로 강한 건열저항성을 가진 자원들이 속하였다. 분 류군별 건열처리 후 최종발아율간의 표준편차는 Ⅰ군에서 10.2로 가장 크게 나타났으며, Ⅱ군, Ⅲ군 및 Ⅳ군은 각각 9.0, 4.9, 3.6으로 건열저항성이 큰 품종군일수록 최종발아 율간 표준편차는 작아지는 경향을 보였다.
Table 5.
Groups of rice germplasm using quartile of final germination after dry-heat treatment (90°C, 36 hours). The values of each quartile were 38% (1st quartile), 65% (2nd quartile) and 85% (3rd quartile).
| Group | Range of germination (%) | No. of accessions | Germination (%) |
|---|---|---|---|
| I | 3~38 | 27 | 22±10.2a† |
| II | 39~65 | 27 | 54±9.0b |
| III | 66~85 | 25 | 78±4.9c |
| IV | 86~98 | 27 | 92±3.6d |
건열처리 후의 발아율에 따라 나눈 4개의 분류군의 벼 생 태형 구분별 자원의 분포는 Fig. 5에서 보는 바와 같다. 인 디카형 벼는 Ⅰ군에 전체의 4.0%가 분포하였으며, Ⅱ군, Ⅲ 군 및 Ⅳ군에 각각 8.0%, 24.0% 및 64.0%가 분포하여, 건 열저항성이 강한 Ⅳ군에 주로 분포함을 알 수 있었다. 이에 반해 자포니카형 벼는 Ⅰ군에 전체의 52.1%가 분포하였으 며, Ⅱ군, Ⅲ군 및 Ⅳ군에 각각 31.3%, 16.7% 및 0%가 분 포하여 인디카형 벼와는 대조적인 분포를 나타내었다. 자바 니카형 벼는 Ⅰ군에 전체의 4.3%가 분포하였으며, Ⅱ군, Ⅲ 군 및 Ⅳ군에 각각 39.1%, 34.8% 및 21.7%가 분포하여, Ⅱ 군과 Ⅲ군에 주로 분포함을 알 수 있었다. 통일형 벼의 경 우 Ⅰ군에는 분포하지 않았으며, Ⅱ군, Ⅲ군 및 Ⅳ군에 각 각 10.0%, 30.0% 및 60.0%가 분포하여 인디카형 벼와 유 사한 건열저항성을 보여주었다.
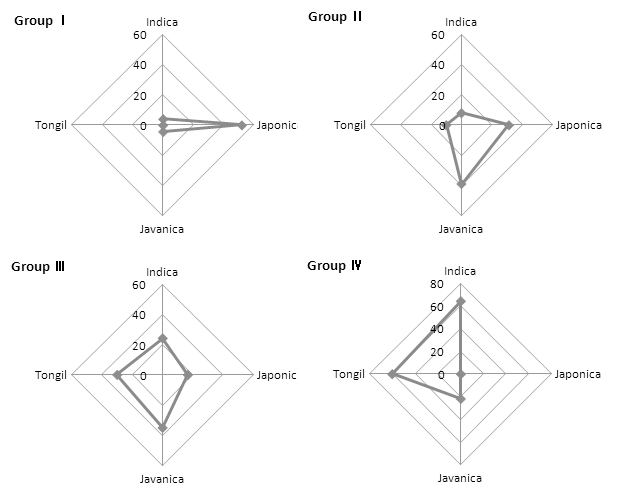
Fig. 5.
Groups of rice germplasm based on germination percentage after dry-heat treatment (90°C, 36 hours). Groups divided by quartile of final germination; GroupⅠ(3~38%), GroupⅡ(39~65%), GroupⅢ(66~85%), GroupⅣ(86~98%).건열처리 후 발아율에 따른 분류군의 생태형별 분포(Fig. 5)는 4°C 저장고 26.5년 보존 후의 발아율에 따른 분류군의 생태형별 분포(Fig. 6)와 유사하므로 건열저항성이 종자수 명과 고도의 유의한 정의 상관관계가 있음(Lee et al., 2006) 을 확인할 수 있었다.
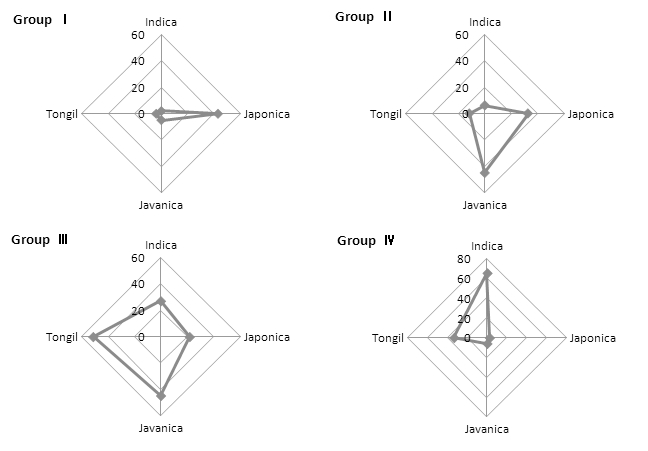
Fig. 6.
Groups of rice germplasm based on germination percentage of seeds stored at 4°C, 30% RH for 26.5 years. Groups divided by quartile of final germination; GroupⅠ(0~12%), GroupⅡ(13~42%), GroupⅢ(43~86%), GroupⅣ(87~100%).건열처리 후 발아율을 대상으로 사분위수를 이용하여 4 개의 분류군으로 나누고(Table 5) 각 분류군의 평균발아율 에 대한 회귀방정식을 구하였다(Fig. 7). 여기서 구한 회귀 방정식을 이용하여 각 분류군별 평균 종자수명을 산출하였 다(Table 6). 건열처리 시 벼 분류군별 종자수명은 Ⅰ군에 속 한 벼의 경우 20시간이며, Ⅱ군, Ⅲ군, Ⅳ군에 속한 벼의 종 자수명은 각각 39시간, 100시간, 411시간으로 계산되었다.
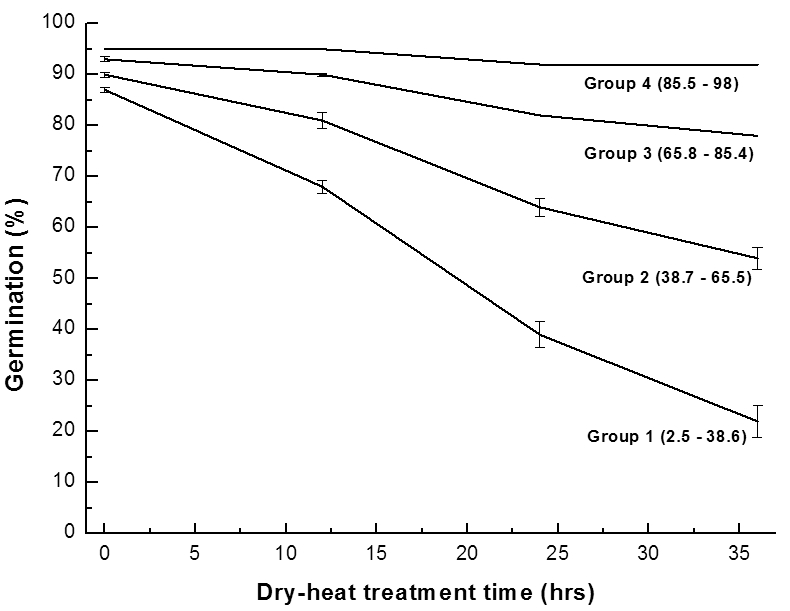
Fig. 7.
Change of germination percentage of rice germplasm accessions grouped after dry-heat treatment (90°C). Vertical bars represent standard errors of the means.Table 6.
Estimating seed longevity (P50) and seed germination percentage of rice germplasm accessions through dryheat treatment (90°C) using multiple regression analysis.
| Group | Dry-heat treatment time (hours) | P50 (hours) | |||
|---|---|---|---|---|---|
| 0 | 12 | 24 | 36 | ||
| I | 87±7.9a† | 68±13.7d | 39±14.7d | 22±10.4d | 20 |
| II | 89±6.0a | 81±7.9c | 64±12.1c | 54±9.2c | 39 |
| III | 93±4.0a | 90±4.6b | 82±6.0b | 78±5.4b | 100 |
| IV | 95±3.5a | 95±3.2a | 92±4.0a | 92±3.6a | 411 |
건열처리 12시간, 24시간 및 36시간 경과 시 분류군별 활 력저하의 정도는 고도의 유의한 수준(p<0.0001)으로 분류 군간 차이를 보였다(Table 6). 건열처리에 따른 분류군별 활 력저하의 경향도 4°C 저장고에 보존 되었던 벼 유전자원의 분류군별 종자수명(Na et al, 2014)과 같은 경향을 보였다.
앞서 4°C 저장고 보존자원의 26.5년 저장 후 발아율에 따 른 분류군 구분(Fig. 6)과 건열처리 36시간 경과 후의 발아 율에 따른 분류군 구분 시 각 분류군에 속하는 벼의 생태형 별 분포 비율(Fig. 5)이 유사함을 알 수 있었다. 그러므로 Table 5를 연결하여 보면 건열처리 36시간 후의 발아율에 따라 구분 된 분류군 중 Ⅰ군에 속하는 자원들은 종자수명 이 21년 정도이며, Ⅱ군, Ⅲ군, Ⅳ군에 속한 자원들은 각각 종자수명이 24, 30, 76년 정도가 될 것으로 예측할 수 있겠 다(Table 7).
Table 7.
Estimating seed longevity (P50) and optimal time (P85) for monitoring of viability in seeds stored at 4°C, 30% RH through dry-heat treatment (90°C, 36 hours).
| Group | Germination percentage after dry-heat treatment at 90°C for 36 hours | Storage at 4°C | |
|---|---|---|---|
| P50 (years) | P85 (years) | ||
| I | 3~38 | 21.1 | 14.8 |
| II | 39~65 | 23.6 | 17.5 |
| III | 66~85 | 30.0 | 20.7 |
| IV | 86~98 | 75.7 | 45.6 |
갱신 된 벼 유전자원을 4°C 저장고 보존 시 효율적인 첫 활력모니터링 시점은 Fig. 7의 회귀방정식을 이용하여 발아 율이 85%로 저하되는 시점(P85)으로 계산해 보면 건열처리 36시간에 따른 발아율이 Ⅰ군에 속하는 자원은 저장 후 14 년, Ⅱ군, Ⅲ군, Ⅳ군에 속한 자원들은 각각 저장 후 17, 20, 45년을 기준으로 하여 설정할 수 있겠다(Table 7).
적 요
벼 유전자원의 효율적인 보존관리를 위해 종자수명 예측 방법을 규명하고자 본 시험을 수행하였다. 효과적인 종자수 명 예측 방법을 규명하기 위해 전년도에 수확한 벼 106품종 을 대상으로 인위노화처리 방법인 노화촉진(AA)처리, 퇴화조 절(CD) 및 건열처리(DHT)를 실시하고, 4°C 저장고에 26.5 년간 보존된 벼 유전자원 3,066점의 종자수명 자료와 비교 분석한 결과는 다음과 같다.
벼 유전자원의 효과적인 종자수명 예측 방법으로는 건열 처리(90°C, 36시간)였다. 전년도 수확한 벼 품종의 건열처 리 후 발아율 성적을 사분위수로 4개의 분류군으로 나누었 을 때, 분류군별로 분포하는 벼 생태형별 품종 비율이 4°C 저장고 보존자원의 최종발아율에 따른 4개 분류군의 분포 비와 흡사하였다. 갱신 된 벼 유전자원을 4°C 저장고 보존 시 효율적인 첫 활력모니터링 시점은 4개 분류군 중 Ⅰ군에 속하는 자원은 저장 후 14년, Ⅱ군, Ⅲ군, Ⅳ군에 속한 자원 들은 각각 저장 후 17, 20, 45년을 기준으로 하여 설정할 수 있겠다. 건열처리는 벼 유전자원 종자수명 예측뿐만 아니라 종자은행에서 보존자원의 효율적인 활력검정 주기 설정 및 갱신 주기 결정에도 도움이 되겠다.
