

최근 온실가스 배출에 의한 지구 온난화 위기와 불안정한 국제 원유가 등의 장기적인 문제해결을 위하여 바이오에너 지가 주목받고 있다. 바이오에너지 원료작물에는 억새, 스 위치그래스 등 초본류와 포플러, 버드나무 등 목본류가 잘 알려져 있다. 그중에서 억새(Miscanthus)는 화본과(Poacea), 기장아과(Panicoideae), 쇠풀족(Andropogoneae) 억새속에 속 하는 식물이다(Chen & Stephen, 2006). 또한 C4 광합성 경 로를 가지는 다년생 식물로서 최근에는 대표적인 섬유질계 바 이오에너지작물 중의 하나로 관심이 증가되고 있다(Atkinson, 2009; Greef et al., 1997; Lewandowski et al., 2000; Yan et al., 2012). 억새속 식물에는 17개 종이 존재한다고 알려 져 있는데 이 중 바이오에너지용으로 중요한 종은 한국 등 동아 시아가 원산인 참억새(M. sinensis)와 물억새(M. sacchariflorus) 그리고 물억새 4배체와 참억새 2배체의 종간 교잡종인 이 질 3배체 억새(M. x giganteous) 등이다(Lewandowski et al., 2000). 억새는 에너지의 투입/생산 수지가 높으면서도 친환 경 재배가 가능한 다년생 작물로 알려져 일찍이 유럽과 북 미 등 선진국에서 재배를 시작하였다(Bullard & Metcalfe 2001). 억새 원산지가 아닌 유럽과 북미에서는 종자가 맺히 지 않아 생태계 교란 우려가 없으면서 바이오매스 수량이 많은 이질 3배체 억새를 재배하고 있다(Moon et al., 2010).
억새 양분 저장기관은 지하 10~20 cm 범위에 분포하는 지 하경이라고 알려져 있다(Chen & Stephen 2006; Lewandowski et al., 2000). Christian et al. (2008)에 의하면 14년간 지속 적인 억새 재배에 질소질 비료 등의 시용량과 무관하게 수 확한 바이오매스 생산량의 차이는 없었다고 보고하고 있으 며, 이는 억새의 생육이 시작되며 지하경에 저장된 양분이 지상부 줄기로 이행하고, 억새의 생육말기인 가을경부터 고사가 될 때까지 지상부에서 지하경으로 이행․저장되어 이 듬해 재활용되기 때문이다(Beale et al., 1996; Himken et al., 1997). 억새는 12월 수확에 비해 2월 수확에서 하위엽 탈락 등으로 인해 수량이 14~15% 감소하고, 3월 수확하면 2월 수확에 비해 13% 감소하지만 바이오매스 중의 회분, 질소, 염소 유황 함량이 감소하여 연료펠릿의 품질이 좋아 진다(Lewandowski & Heinz 2003).
억새 수확작업은 애취, 집초, 곤포 및 수거 등의 과정을 거치는데 보통 트랙터 동력을 활용하여 수행한다. Michael et al. (2015)은 3년 동안 맹아 전, 후 트랙터 답압으로 인한 토양경도, 억새 생육반응을 조사하였는데 무처리의 11.17 톤/ha에 비해 매년 트랙터 이용 곤포 + 수거로 인한 답압구 에서 바이오매스 수량이 5.24 톤/ha로 가장 낮았고 토양 경 도도 2.74 Mpa에서 3.08 MPa로 유의적으로 증가하였다. 트랙터 이용 억새 바이오매스 수확 시 수확시기가 지연될 수록 토양경도와 진비중이 유의적으로 증가하였고 수분 침 투계수는 유의적으로 낮아졌다. 특히 맹아 4주 후 수확하면 경수와 경장 등 생육량이 적어 바이오매스 수량이 크게 감 소하였다(O’Flynn et al., 2011). 더 나아가 트랙터 답압으 로 인한 억새 생육에 미치는 부작용을 감소시키기 위한 연 구도 수행되고 있다. 그 중에서 억새 수확 시 타이어 공기 압을 줄임으로서 토양경도 등 물리성 악화와 지하경 손상 을 감소시킴으로서 이듬해 수량 감소를 최소화 할 수 있었 다(Michael et al., 2014).
국내에서 억새는 작물로 이용된 역사가 짧기 때문에 재 식, 포장관리, 수확 등 기본적인 재배기술이 확립되지 않아 고 수량성 등 억새 특유의 장점이 발휘되지 않은 실정이다. 특히 억새 수확 한계기가 정립되지 않아 늦은 수확 시 트랙 터 등 중장비의 답압으로 인해 양분 저장기관인 지하경이 손상되어 생육에 큰 지장을 초래할 것으로 예상된다. 따라 서 본 연구는 에너지용으로 재배되는 ‘거대1호’, ‘도입 이 질 3배체’ 의 수확 지연에 따른 출수율과 생육량 감소 양상 을 조사하여 작물로서 억새의 수확 한계기를 정립하기 위 하여 수행하였다.
재료 및 방법
시험포장 및 식물재료
본 시험은 전북 익산시 용안면 석동리 소재 ‘거대억새 생 산단지’(36°14′34.55″N, 126°93′77.95″E)에 조성된 재식 5 년차의 억새 품종비교 전시포에서 수행하였다. 시험 포장 은 억새밭 조성 이전에 논으로 경작되었던 해안평탄지로서 지하 30 cm까지 미사질 양토, 그 이하에서는 사질토양 이 었다. 시험에 사용된 억새 품종은 국립식량과학원에서 육 성한 물억새(M. sacchariflorus)의 일종인 ‘거대1호’와 미국 일리노이즈대학교에서 도입한 이질3배체(M. x giganteous) 였다.
억새 수확 및 수확시기 설정
억새 수확 트랙터 동력 작업은 예취(Mow & conditioning), 곤포(Baling), 곤포 수집(Collecting of bales) 등 3단계를 거 치는데 이 중 대형 장비가 동원되는 단계는 곤포, 곤포수집 단계이다(O’Flynn et al., 2015). 트랙터는 4.6톤, 280마력 (FENDT 828, AGCO GmbH, Germany) 기종을 사용하였 으며, 앞바퀴와 뒷바퀴에 장착된 타이어 규격은 각각 600/70 R30, 710/70 R42s였다. 트랙터에 장착된 곤포기(RP 535/545, Lely, Netherland)는 크기 3.39×2.04×2.085 (m), 무게 2.25 톤이었다.
수확시기는 3 시기로 설정하였으며, 지하경에서 맹아가 시작되기 전으로 ‘맹아전’으로 판단되는 4월 1일, m2 당 새 싹이 20개 이상 출현하여 보라색 비늘잎이 보여 ‘맹아기’로 판단되는 4월 10일, 그리고 m2 당 새싹이 40개 이상 출현 하고 어린 잎이 전개되어 ‘맹아종’으로 판단되는 4월 17일 로 하였다(Fig. 1). 수확작업은 Michael et al. (2015)이 실 시하였던 방법으로 3월 하순경 억새 줄기를 인력 수확 후 상기 기종의 트랙터와 곤포기를 연결하여 수확 시기별로 시험구를 1~2 km/h 속도로 5회 답압하였다. 수확 전, 후(3 월 상순~4월 하순) 기상자료는 시험포장에서 21 km 거리 의 전라북도 군산시 내흥동 소재 군산기상대 관측 평균기 온, 지표온도 및 강수량을 활용하였다.

토양경도 및 억새 생육특성 조사
토양경도는 억새의 생육이 완료되고 고사되는 시기에 산 중식 경도계(Yamanaka, Japan)를 이용하여 지표면으로부터 지하 10 cm와 20 cm 깊이의 토양경도를 측정한 후 절대 경도를 아래의 식으로 산출하였다.
시험구의 경장과 경수의 생육시기별 변화를 알기 위해 생육초기(4월 29일)부터 잎과 줄기 고사기(10월 30일)까지 4차례 조사하였다. 수확 직후 각 시험구별로 3 지점씩 1×1 m 크기의 격자를 설치하여 매 조사시기별 동일 지점을 조사 하였는데 경장은 줄기의 지제부에서 최 상위엽 끝까지 길 이를, 경수는 격자내 줄기 중 최 상위엽이 고사한 줄기를 제외한 완전한 줄기의 수를 조사하였다. 경장, 경태, 경수 등 수량 구성요소와 마디수 및 출수율은 잎과 줄기가 고사 한 10월 30일에 각 처리별로 3반복씩 예취한 후 조사하였 고, 수량은 전남 무안에 위치한 국립식량과학원 바이오에 너지작물연구소의 건조기에 80°C 온도로 3일동안 건조 후 조사하였다. 2014년 및 2015년 생성 지하경태는 10월 30일 수량조사를 위한 줄기를 예취한 각 시험구별로 지하경의 굵기를 조사하였다. 물억새 줄기는 전년도 생성 지하경 정 단부가 맹아하여 자라고, 당년에는 줄기의 지하부 마디 눈에 서 새롭게 지하경이 생성되어 신장하고 육안으로 정단부를 확인할 수 있다(Chen & Stephen 2006). 따라서 본 연구에서 는 줄기를 중심으로 정단부가 있는 쪽의 지하경을 당년도 생 성으로, 그 반대 쪽을 전년 생성 지하경으로 판단하였다.
결과 및 고찰
전년도(2014년도) 생육 바이오에너지용 억새의 늦은 수 확에 따른 생육 완료 후 시험포장의 깊이별 토양경도 조사 결과는 Fig. 2와 같다. 지하 10 cm 토양 경도는 맹아시(4월 1일), 맹아기(4월 10일), 맹아종(4월 17일) 수확에서 각각 650, 793, 735 kPa로 일정한 경향이 없었으나, 지하 20 cm 에서는 각각 741, 915, 1,045 kPa로 나타나 수확이 지연될 수록 지하 20 cm의 토양경도가 높다는 것을 알 수 있었다. 억새는 영년생 작물로 깊이갈이 등 토양 배양작업은 재식 당년에만 실시할 수 있다(Moon et al., 2010). 늦게 수확한 시험구일수록 생육완료 후인 10월 하순에 지하 20 cm 토 양경도가 높았던 것은 답압과 Table 1에서와 같이 4월 상․ 중순에 평균기온과 지표온도가 10°C 이상으로 높고, 강수 량이 각각 38.0 mm, 30.6 mm로 많아 땅이 마르지 않은 상 태에서 트랙터와 곤포기에 의한 답압의 영향인 것으로 판 단된다. 억새의 양분 저장기관인 지하경은 지하 10 cm 부 근에 분포하는데 저장양분이 봄에는 지상부로 이행하여 영 양생장을 하고 가을철 잎과 줄기가 시듦에 따라 다시 지하 부로 이행하여 새롭게 생성된 지하경에 저장된다(Himken et al., 1997). 지하 10 cm 토양 경도는 수확 시기별로 차이 가 크지 않았는데 이것은 4월 중순 트랙터와 곤포기 답압 으로 인해 단단해진 토양이 새로 생성된 지하경 신장으로 개선되었기 때문인 것으로 생각된다.
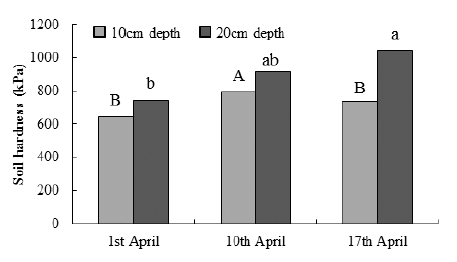
Fig. 2
Soil penetration resistances at 10-cm and 20-cm depth in the growth completion stage of Miscanthus at each delayed harvesting date. Means with the same letter are not significantly different at p < 0.05.Table 1
Climate data of 2015 before and after the harvest of Miscanthus in the experimental fields.
전년도 생육 바이오에너지용 억새의 늦은 수확 시기별 당년 출현경수의 경시적 변화를 조사한 결과는 Fig. 3과 같 다. 거대1호, 이질 3배체 모두 처리내용에 관계없이 경수는 생육 중기에 최대에 달하였고, 그 후 생육시기가 경과함에 따라 줄어드는 경향이었다. 거대 1호는 생육 중기(5월 22 일)에 맹아시 수확에서 경수가 122개/m2 , 맹아종 수확에서 169개/m2로 수확 시기에 따라 차이가 컸으나 생육 최성기 (7월 22일)에 각각 62개/m2 , 70개/m2로 급격히 감소하였고 그 차이도 적었다. 이질3배체는 생육 중기(5월 22일)에 맹 아시 수확 101개/m2 , 맹아종 수확 114개/m2로 그 차이가 적었고 생육 최성기(7월 22일)에 각각 65개/m2 , 74개/m2로 감소폭도 적었다. 거대1호는 물억새(M. sacchariflorus)의 일종으로 지하경의 절간장이 10~15 mm 정도로 길고 직선 형으로 길게 신장하지만 이질 3배체 억새(M. x giganteous) 는 절간장이 5 mm 이하로 짧고 만곡형으로 물억새에 비해 짧게 신장하는 특성이 있다(Moon et al., 2010). 억새의 지 하경은 문자 그대로 일종의 줄기이기 때문에 정단우세성으 로 인해 지하경이 손상되지 않은 정상적인 조건에서는 정 단부가 아닌 마디의 눈에서 신초가 발생하지 않지만 정단 이 손상되면 아래쪽 마디의 눈에서 신초가 발생한다(Morris, 1991). 맹아종 수확 등 늦게 수확한 구에서 생육 중기에 거 대1호의 경수가 많은 것은 정단이 손상된 지하경이 많았고 지하경의 절간장이 길어 마디의 눈에서 발생한 신초 수가 많았지만 이질3배체는 손상된 지하경이 많았을지라도 지하 경 절간장이 짧아 신초 발생이 적었기 때문에 경수가 적은 것으로 생각된다.

Fig. 3
Sequential changes in stem number in juvenile (April 29), mid (May 27), luxuriant (July 22), and senescence (October 30) stages of Miscanthus sacchariflorus cv. Geodae 1 (a) and Miscanthus × giganteus (b) at each delayed harvesting date.전년도 생육 바이오에너지용 억새 포장의 늦은 수확 시 기별 당년 신장 줄기의 생육 완료 후 전년(2014년) 및 당년 (2015년) 생성 지하경을 굴취하여 굵기를 조사한 결과는 Fig. 4와 같다. 맹아시 수확에서 전년 및 당년 지하경태가 거대 1호는 각각 8.5 mm, 8.4 mm로 거의 차이가 없었고 이질 3배체 억새는 각각 8.4 mm, 8.7 mm로 오히려 약간 굵어졌다. 그러나 늦게 수확하면 거대 1호는 전년 및 당년 지하경태가 맹아기 수확에서 각각 8.3 mm, 7.5 mm 맹아종 수확에서 각각 9.2 mm, 6.8 mm로 가늘어졌다. 이질3배체 또한 거대 1호에 비해 그 감소폭이 적었으나, 전년 및 당년 지하경태가 맹아기 수확에서 각각 8.4 mm, 8.2 mm, 맹아 종 수확에서 각각 8.5 mm, 8.0 mm로 가늘어진 경향이었 다. 억새의 양분 저장기관은 지하경으로서 봄과 여름에는 전년도 저장된 지하경의 양분이 지상부로 이동하여 생육하 고 가을에 식물체가 노화됨에 따라 지상부 양분이 다시 당 년 생성된 지하경으로 이동하여 저장된다(Beale et al., 1996; Christian et al., 2008). 늦게 수확할수록 당년 생성된 지하 경이 가늘어지는 것은 늦은 수확에 따른 지상부 생육이 부 진하여 가을철 지하경으로 이동될 양분이 상대적으로 적었 기 때문인 것으로 추측되지만 금후 세부적인 연구와 검토 가 필요할 것으로 생각된다. 아울러 당년 생성된 지하경이 차년도 지상부 생육에 크게 영향을 끼치므로(Beale et al., 1996; Christian et al., 2008) 늦게 수확하여 지하경이 가늘 어진 곳에서는 차년도 생육이 부진할 것으로 예상된다.

Fig. 4
Rhizome diameters of (a) Miscanthus sacchariflorus cv. Geodae 1 and (b) Miscanthus × giganteus produced in previous and current years at each delayed harvesting date. Means with the same letter are not significantly different at p > 0.05.전년도 생육 바이오에너지용 억새 포장의 늦은 수확 시 기별 당년 신장 줄기의 생육 완료 후 경장과 줄기 수량을 조사한 결과는 Fig. 5와 같다. 거대1호, 이질3배체 모두 늦 게 수확할수록 경장이 작아지고 아울러 줄기 수량도 감소 하였다. 거대1호와 이질3배체의 경장은 맹아시 수확에서 각각 308 cm, 374 cm였으나 수확시기가 늦을수록 작아져 맹아종 수확에서는 각각 268 cm, 341 cm로 작아졌다. 줄기 수량 또한 맹아시 수확에서는 각각 1,440 kg/10a, 1,777 kg/10a 이었으나 맹아종 수확에서는 각각 847 kg/10a, 963 kg/10a 로 크게 감소하였다. 억새 줄기 수량은 전년 12월 수확 대 비 2월 수확은 14~15%, 3월 수확은 25~26% 감소하지만 회분, 염소, 유황 등 연료의 품질을 악화시킬 수 있는 성분 도 감소하기 때문에 바이오매스 사용 목적에 따라 수확시 기를 조절할 수 있다(Lewandowski & Heinz, 2003). 그러 나 수확기가 너무 늦어 지하경의 새싹이 트기 시작한 이후 수확하면 트랙터 등 중장비에 의한 답압으로 토양경도 등 물리성이 악화되고 새싹이 손상되어 4월에 중장비를 동원 하여 수확하면 바이오매스 수량이 약 6톤/ha로 동일 시기 의 인력수확 13.9톤/ha에 비해 크게 감소한다(Michael et al., 2015). 종자번식작물인 밀도 트랙터 답압횟수가 7회 정 도로 너무 많으면 토양 물리성이 악화되어 수량이 무 답압 대비 18~38% 감소한다(Tolon-Becerra et al., 2011). 지하 경으로 번식하는 작물인 억새는 그 답압으로 인한 부작용 이 심할 것으로 생각되므로 트랙터 등 중장비 이용 수확시 공기압을 감소시킨 타이어가 장착된 트랙터를 사용하는 (Michael et al., 2014) 등 답압 피해를 감소시킬 수 있는 연 구가 필요할 것으로 생각된다.
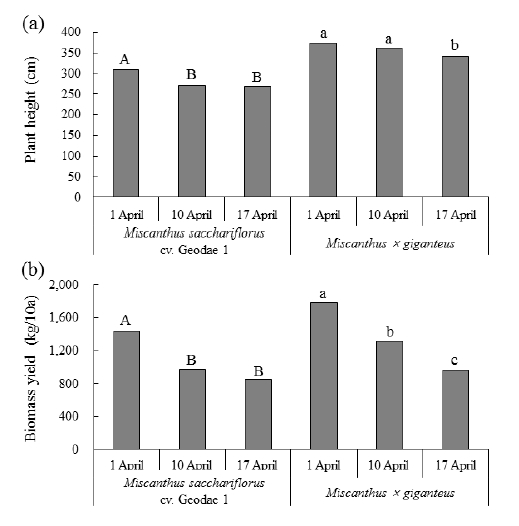
Fig. 5
Plant height (a) and biomass yield (b) in the growth completion stage of Miscanthus sacchariflorus cv. Geodae 1 and Miscanthus × giganteus at each delayed harvesting date. Means with the same letter are not significantly different at p > 0.05.전년도 생육 바이오에너지용 억새 포장의 늦은 수확 시 기별 당년 신장 줄기의 출수율과 줄기의 마디수를 조사한 결과는 Fig. 6과 같다. 거대1호의 경우 출수율이 맹아시 수 확에서 3.9%로 극히 낮았고 맹아기 및 맹아종 수확에서는 거의 출수되지 않았다. 반면에 이질 3배체의 경우 맹아시 수확에서 49.6%로 거대 1호에 비해 월등히 높았고, 수확기 가 맹아기, 맹아종으로 늦어지더라도 출수율이 각각 40.6%, 21.8%로 맹아시에 비해 낮아졌지만 동일 수확시기의 거대 1호에 비해서는 높았다. 줄기의 마디수는 거대 1호의 경우 맹아시, 맹아기 및 맹아종 수확에서 각각 18.5, 17.3, 16.7 개였으나 이질 3배체의 경우 맹아시, 맹아기 수확 모두 17.7개, 맹아종 수확에서 17.3개로 거대 1호에 비해 적었고 수확기가 늦어져도 그 감소폭도 적었다. 벼과식물 출수의 빠르고 늦음은 광주기, 온도 및 기본영양생장성으로 결정 되는데, 벼의 경우 고위도 지방에서는 기본영양생장성과 감광성이 작고 감온성이 큰 품종이 적응하는 반면, 저위도 지방에서는 기본영양생장성과 감광성이 큰 품종이 적응한 다고 하였다(Noh et al., 1997). 본 연구의 시험에 사용된 거대 1호는 국내 각지의 수집지역 중에서 비교적 저위도인 35°N 이하의 남부 지역에서 자생하는 수집종으로서 타 지 역에서 수집한 것에 비해 생육량이 월등히 많다(Moon et al., 2010). An et al. (2015)에 의하면 거대 1호는 맹아일로 부터 지엽전개일까지의 생육일수가 타 억새에 비하여 월등 히 길며 이는 영양생장기간이 길어서 바이오매스 축적양이 많은 것으로 보고하였다. 따라서 거대 1호가 이질3배체에 비해 출수율이 낮으면서 줄기의 마디수가 많은 것은 기본 영양생장성이 상대적으로 크기 때문인 것으로 사료된다.

적 요
바이오에너지용 억새의 트랙터 부착 수확기 등 중장비 동원 늦은 수확에 따른 부작용을 구명하기 위해 전년도 생 육 줄기의 늦은 수확 시기별 당년 생육특성과 마른줄기 수 량을 구명한 결과는 다음과 같다.
1. 시험포장의 지하 10 cm 토양 경도는 맹아시, 맹아기, 맹 아종 수확에서 각각 650, 793, 735 kPa로 일정한 경향이 없었으나 지하 20 cm에서는 각각 741, 915, 1,045 kPa 로 늦게 수확할수록 단단해 졌다.
2. 이질 3배체에 비해 거대1호가 생육시기별 경수의 변 이폭이 컸다. 거대 1호는 수확 시기별로 가장 늦은 맹 아종에서 생육중기(5월 22일)에 169개/m2로 가장 많 았으나 생육 최성기(7월 22일)에 70개/m2로 급격히 감소하였다.
3. 수확시기가 늦을수록 당년 생성된 지하경이 전년의 것에 비해 가늘어져 맹아종 수확에서 전년, 당년생성 지하경태가 거대 1호는 각각 9.2 mm, 6.7 mm 이질 3 배체는 각각 8.5 mm, 8.0 mm였다.
4. 수확시기가 늦을수록 경장이 작아지고 아울러 줄기 수량도 감소하였다. 거대1호와 이질3배체의 경장은 맹아시 수확에서 각각 308 cm, 374 cm였으나 맹아종 수확에서는 각각 268 cm, 341 cm로 작아졌다. 줄기 수 량 또한 맹아시 수확에서는 각각 1,440 kg/10a, 1,777 kg/10a였으나 맹아종 수확에서는 각각 847 kg/10a, 963 kg/10a로 크게 감소하였다.
5. 출수율은 거대 1호의 경우 맹아시 수확에서 3.9%로 극히 낮았고, 맹아기 및 맹아종 수확에서는 거의 출수 되지 않았다. 이질3배체는 맹아시 수확에서 49.6%였고, 맹아기, 맹아종에서도 각각 40.6%, 21.8% 수준을 유 지하였다.
