

서 론
재료 및 방법
품종 선정
재배방법
시료 채취 및 조사방법
분석방법
데이터 분석
통계 처리
결 과
등숙형질 변이
이화학적 특성 변이
Sigmoid equation을 이용한 수량지표 예측
고 찰
적 요
서 론
우리나라의 연간 1인당 쌀 소비량은 다양한 먹을거리의 증가로 인하여 1980년 132.4 kg에서 2000년 93.6 kg, 2019년 59.2 kg으로 점차 감소하고 있다(Statistics Korea, 2020). 이러한 추세에 대응하여 쌀을 밥이나 떡으로만 먹는다는 기존의 인식에서 벗어나 제면, 제빵 등의 다양한 용도로 이용할 수 있도록 가공기술이 개발되어 왔다. 쌀 가공시장의 주요 원료는 쌀가루인데, 이에 따라 쌀가루의 생산성과 품질의 향상을 위한 벼 품종이 육성되어왔다. 최근 육성된 ‘쌀가루 가공용 품종’은 물에 불리지 않아도 종자가 쉽게 부서지는 특성을 지녀 제분공정과정이 축소되고 품질 및 저장성이 향상되는 등의 강점을 가지고 있는 것으로 알려져 있다.
한편 벼에서 등숙기 온도는 수량과 품질에 영향을 미치는 주요한 환경요인이다. 자포니카 품종에서 천립중이 최대가 되는 온도는 보통 22°C 내외로 알려져 있다. Kim (1983)은 천립중이 최대가 되는 온도는 22°C이었지만 16°C의 저온에서도 유의한 감소는 보이지 않았으며 25°C의 고온에서는 약 6%의 감소를 보였다고 하였다. 또한 출수 이후 40일간의 평균온도 22°C 전후에서 현미천립중이 최고를 보였으며, 그 이상에서는 천립중이 감소했다고 하였다(Choi et al., 2011). 등숙률의 경우 등숙기 온도가 22°C 전후에서 최고값을 보였지만 19°C의 저온에서도 2%의 차이에 그쳤다는 기존의 보고가 있으며(Yoshida & Hara, 1977), Park et al. (1999)은 등숙기에 30°C 전후의 고온이 등숙률을 감소시킨다고 하였다. 천립중과 밀접한 관련이 있는 종자 크기(Li et al., 2018)의 경우 입장(종자길이)은 30.5°C 이상에서, 입폭(종자너비)은 27.5°C 이상의 고온에서 감소하며, 고온으로 갈수록 장폭비가 커진다고 하였다(Tashiro & Wardlaw, 1991a).
이와 같이 일반 벼 품종의 등숙기 온도에 따른 등숙형질의 변이에 대한 연구는 오래전부터 수행되어왔다. 최근 쌀가루용 품종이 개발됨에 따라, 육성품종에 맞는 지역별 적합 이앙시기 등의 재배연구가 수행되고 있지만 등숙기 온도에 따른 등숙형질과 이화학적 특성 변화에 대해서는 연구된 바가 없다. 따라서 본 논문에서는 등숙기 온도에 따른 쌀가루용 벼의 등숙특성 및 이화학적 특성 변이에 대하여 구명하여 쌀가루용 품종의 재배기술을 설정하는 데 기초자료로 제시하고자 한다.
재료 및 방법
품종 선정
쌀가루용 품종은 전분구조의 차이로 인하여 찹쌀과 같이 분상질의 불투명한 빛을 띠게 되며, 분질배유의 많고 적음에 따라 적으면 연질미, 많으면 분질미로 분류된다. 이 같이 분류된 품종들의 등숙기 온도에 따른 등숙특성 차이를 알아보고자 재배면적 및 분류품종의 대표성 등을 고려하여 새누리(일반 멥쌀), 설갱(연질미), 바로미2 (분질미)를 선정하였다.
재배방법
본 실험은 2019년에 국립식량과학원 내 위치하며 온도 및 습도 등의 조절이 가능한 인공기상동에서 실시되었다. 모종판(406구, 11*11 mm)에 1립씩 파종하여 14일간 키운 모를 6월 14일에 1/5,000a 와그너포트에 포트 1개당 1본씩 3주를 이앙하였으며, 품종당 50포트씩 총 150포트를 제작하여 출수기까지 노지상태로 재배하였다. 출수기는 이삭마다 출수일자를 모두 표시하였는데, 이삭이 잎집에서 절반쯤 추출되었을 때를 기준으로 하였다. 등숙기 온도처리 전 시료의 균일성을 확보하기 위하여 출수 직전 포트별로 생육정도를 조사하고 각 그룹을 비슷한 수준으로 맞추어 주었으며, 그 평균값은 Table 1과 같다. 또한 출수 직후의 온도변화는 임실률에 영향을 미칠 수 있기 때문에 출수기 약 3일 후에 각각 온도가 다른 3개의 growth chamber로 나누어 이동하였다. 각 chamber의 온도는 저온부터 고온까지 평균 18°C (23/13°C), 22°C (27/17°C), 28°C (33/23°C)로 설정하였다. Chamber의 광조건은 자연상태와 동일하며 상대습도는 70% 정도로 유지하였다.
Table 1.
Growth at the heading date before setting temperatures.
| Varieties |
Temperature (°C, day/night) | Mean Temp. | Growth | ||
| Plant Height (cm) | no. of tillers per plant | no. of plants | |||
| Saenuri | 23/13 | 18°C | 90.5 | 10.8 | 51 |
| 27/17 | 22°C | 90.5 | 11.0 | 48 | |
| 33/23 | 28°C | 90.8 | 11.4 | 51 | |
| ns | ns | ||||
| Seolgaeng | 23/13 | 18°C | 82.6 | 11.0 | 51 |
| 27/17 | 22°C | 83.1 | 10.5 | 48 | |
| 33/23 | 28°C | 82.2 | 11.1 | 51 | |
| ns | ns | ||||
| Baromi2 | 23/13 | 18°C | 81.3 | 9.0 | 51 |
| 27/17 | 22°C | 80.6 | 9.2 | 48 | |
| 33/23 | 28°C | 81.6 | 8.9 | 51 | |
| ns | ns | ||||
시료 채취 및 조사방법
종실은 1주당 5개의 이삭에서만 채취하였다. 이삭별 출수일에 따라 출수 후 적산온도 200°C부터 1200°C까지 총 7회 채취하였고 채취 직후 등숙립과 비립의 구분 없이 정조천립중을 조사하였다. 적산온도 1200°C에 이른 수확용 시료는 채취하여 등숙률, 현미천립중, 종자크기 등을 조사한 뒤 동결건조 시켰다가 현미상태로 분쇄하고 전분함량, 단백질함량 등을 분석하였다. 조사방법은 농업과학기술 연구조사분석기준에 준하였다.
분석방법
전분함량은 변형된 AOAC Official Method 996.11 방법으로 Total Starch kit (megazyme International Ireland Ltd., Ireland)를 이용하여 분석하였으며 단백질함량은 원소분석기(Elementar Analyzer System, Vario Macro, Germany)를 이용하여 총 질소함량을 구하고 이 값에 보정계수 5.95를 곱하여 계산하였다.
데이터 분석
Sigmoid Equation은 시간에 따른 종실중 등의 수량 관련 값과 잘 부합하는 것으로 알려져있다(Sheehy et al., 2004). 따라서 등숙 기간에 따른 정조천립중의 변화를 Sigmoid Equation에 적합시켰다. 본 연구에서 사용한 식 (1)은 아래와 같다.
식 (1)에서 t는 출수이후 유효적산온도(Threshold temperature: 8°C), W는 정조천립중이다. a는 최종 천립중이며, t0는 W값이 최종 천립중의 절반(1/2a)이 될 때이자 등숙속도(dW/dt)가 최대가 되는 때의 유효적산온도이다. t=t0일 때의 최대 등숙속도는 a/4b와 같다. a, b, t0와 같은 계수들은 Sigmaplot 10.0 (Systat)을 통해 추정되었다.
통계 처리
모든 실험은 3번 이상 반복하였으며 평균치를 결과 값으로 나타내었다. 본 연구에서의 통계분석은 R (version 4.0.3, R Foundation, Austria)을 이용하여 분산분석 후 Duncan’s Multiple Range Test로 P < 0.05 수준에서 유의적인 차이를 검정하였다.
결 과
등숙형질 변이
등숙기 온도조건에 따른 품종별 등숙률, 현미천립중 및 현미 크기의 변화는 Table 2와 같다. 등숙률은 세 품종 모두 모든 품종에서 22°C일 때 최고치를 보였으며 저온(18°C)와 고온(28°C)조건에서는 감소하는 경향을 보였지만, 모든 품종에서 온도에 따른 유의한 차이는 발생하지 않았다.
Table 2.
Yield components and grain sizes of brown rice in three different varieties under different temperature conditions.
| Varieties |
Temp. (°C, day/night) | Mean Temp. | Yield components | Grain size | ||
| Rate of grain filling (%) | 1,000 Grain weight (g) | Length (mm) | Width (mm) | |||
| Saenuri | 23/13 | 18°C | 78.7 | 22.1a | 5.03 | 2.93 |
| 27/17 | 22°C | 79.6 | 21.1b | 5.02 | 2.86 | |
| 33/23 | 28°C | 76.4 | 19.9c | 4.93 | 2.82 | |
| ns | ** | ns | ns | |||
| Seolgaeng | 23/13 | 18°C | 71.2 | 20.4a | 4.93 | 2.89 |
| 27/17 | 22°C | 73.3 | 20.2a | 4.90 | 2.85 | |
| 33/23 | 28°C | 73.2 | 17.9b | 4.87 | 2.80 | |
| ns | ** | ns | ns | |||
| Baromi2 | 23/13 | 18°C | 79.4 | 21.0a | 4.88 | 2.97a |
| 27/17 | 22°C | 80.8 | 19.5b | 4.84 | 2.92a | |
| 33/23 | 28°C | 79.1 | 15.3c | 4.78 | 2.87b | |
| ns | ** | ns | * | |||
현미천립중의 경우 온도조건과 품종에 따라 매우 유의한 차이를 보였다. 모든 품종에서 온도가 상승할수록 현미천립중은 감소하는 경향을 보였다. 평균온도 22°C를 기준으로 저온 조건(평균온도 18°C)에서는 새누리가 약 4%, 설갱이 약 1%, 바로미2는 약 8%의 증가를 보였으며 새누리와 바로미2는 통계적으로도 유의한 차이가 있었다. 고온 조건(평균온도 28°C)에서는 품종간 큰 차이를 보였다. 모든 품종에서 큰 감소를 보였고 통계적으로도 매우 유의한 차이가 있었다. 다만 감소폭은 품종에 따라 차이가 있었다. 평균온도 22°C와 대비하였을 때 새누리는 21.1 g에서 19.9 g으로 약 6%가 감소했지만, 반면 설갱은 2배인 약 12%, 바로미2는 약 21%로 3배 이상의 감소폭을 보였다.
온도에 따른 현미 크기는 현미 입장(grain length)은 모두 유의한 차이는 나타나지 않았지만 온도가 높아질수록 조금씩 줄어드는 경향을 보였다. 입폭(grain width)의 경우도 입장과 같이 모든 품종이 온도가 높아질수록 감소하는 경향이었는데, 분질미인 바로미2가 고온조건(28°C)에서 약 2% 감소하였으며 통계적으로도 유의한 차이를 보였다.
이화학적 특성 변이
Table 3는 온도에 따른 벼 종실의 이화학적 특성의 변화를 나타낸 결과이다. 전분의 상대 함량은 품종마다 조금씩 차이가 있었다. 일반과 쌀가루용 품종 모두에서 평균온도 18°C와 22°C가 비슷했고 고온조건(28°C)에서는 감소하였다. 그 중에서도 특히 바로미2에서 고온조건일 때 10% 이상 감소하면서 전분 축적이 현저히 줄어든 것을 확인할 수 있었다.
Table 3.
Physicochemical properties of the different varieties under different temperature conditions.
| Varieties |
Temp. (°C, day/night) | Mean Temp. | Total Starch (%) | Protein (%) |
| Saenuri | 23/13 | 18°C | 84.1 | 7.22c |
| 27/17 | 22°C | 84.9 | 7.56b | |
| 33/23 | 28°C | 81.6 | 7.95a | |
| ns | ** | |||
| Seolgaeng | 23/13 | 18°C | 79.4 | 7.06c |
| 27/17 | 22°C | 78.8 | 7.46b | |
| 33/23 | 28°C | 76.5 | 7.82a | |
| ns | ** | |||
| Baromi2 | 23/13 | 18°C | 74.5 | 6.93b |
| 27/17 | 22°C | 77.6 | 6.83b | |
| 33/23 | 28°C | 67.4 | 8.58a | |
| ns | ** |
단백질 함량은 등숙기 온도의 상승에 따라 유의하게 증가하였다. 고온(28°C)에서는 모든 품종에서 고도로 유의한 차이를 보이며 증가하였는데, 22°C와 대비했을 때 새누리와 설갱이 0.3∼0.4% 증가하는 것에 그친 반면, 분질미인 바로미2는 약 1.7%가 증가하며 품종 중에서 가장 높은 증가를 보였다.
Sigmoid equation을 이용한 수량지표 예측
유효적산온도(Effective accumulated temperature)란 식물의 생육단계 등을 예측할 수 있도록 고안된 것으로 평균기온에서 임계온도(Threshold temperature)를 뺀 것을 적산한 값이다. 벼가 등숙을 멈추게 되는 최저 임계온도는 등숙단계마다 다르며, 낮게는 3.4°C부터 높게는 14.2°C에 이르는 다양한 주장이 있지만 보통 8°C~10°C 사이로 알려져 있다(Counce et al., 2009; Ebata, 1990; Gao et al., 1986; Matthews et al., 1995). 본 시험에서 여러 임계온도를 식 (1)의 함수에 적합시켜 본 결과 그래프가 가장 잘 설명되는 온도는 8°C였다. 임계온도를 8°C로 설정하여 유효적산온도에 따른 정조천립중 변화를 sigmoid 함수로 표현한 것을 품종별로 Fig. 1의 그래프로 나타내었고 이 때 함수의 계수(파라미터)는 Table 4와 같다. 유효적산온도는 실제 생육에 관여된 온도에 식물체가 반응하는 기간을 의미하므로 하나의 품종은 온도에 관계없이 하나의 함수로 표현될 수 있다(Lee et al., 2009). 새누리와 설갱이 0.95, 0.93 이상의 결정계수(R2)를 보이며 유효적산온도에 따른 정조천립중의 변화가 잘 설명되었던 것과 달리 바로미2는 약 0.79에 그치는 결과를 보였다. 바로미2의 낮은 결정계수는 저온(18°C)처리구의 이상치에 기인한 것으로 판단되어 저온에서의 값을 떼어 내어 각각 분석하였고, 그 결과 0.97, 0.98 이상의 결정계수 값이 도출되었다.
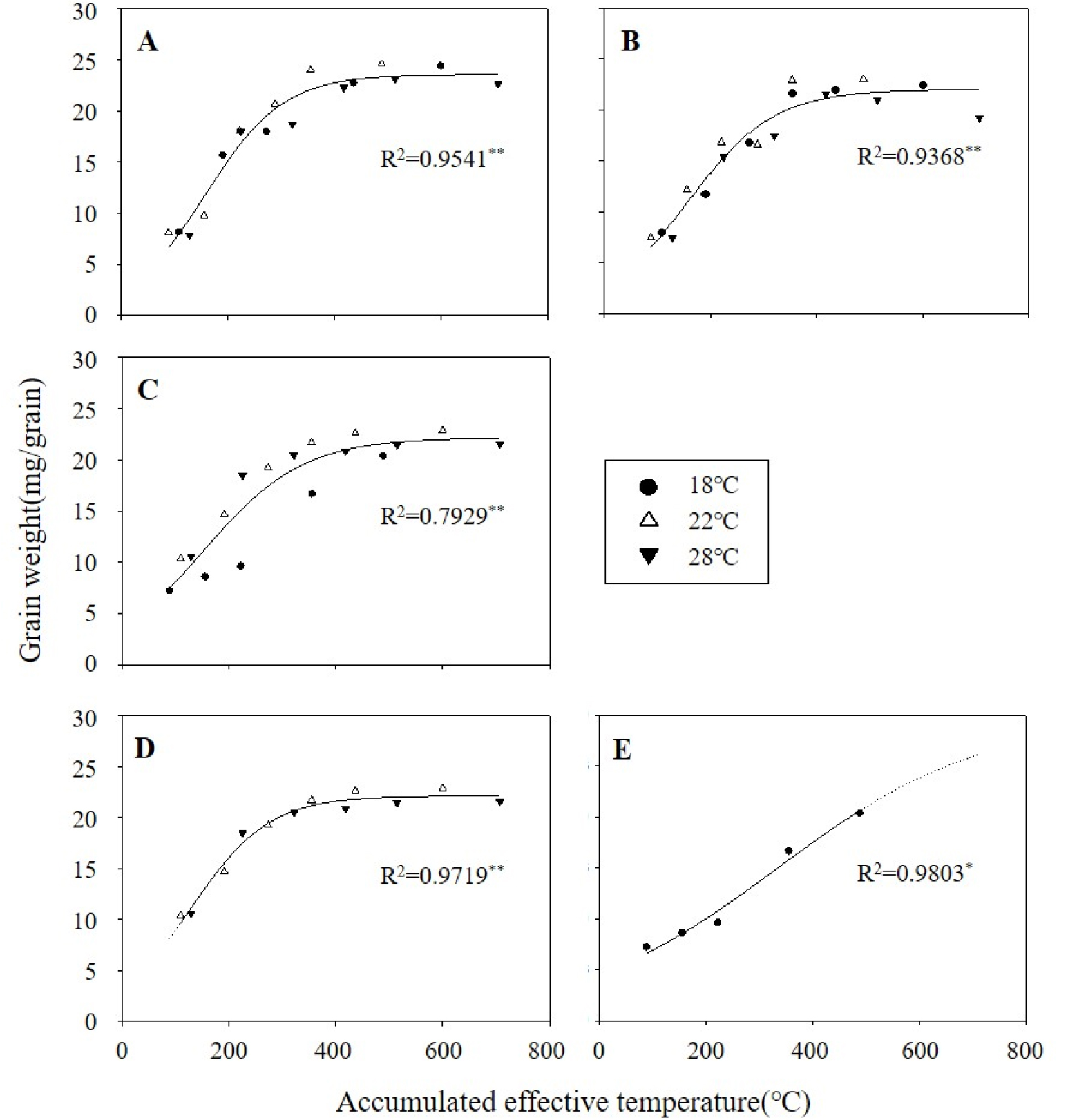
Fig. 1.
Change of grain weights of rough rice in different varieties. The curves were fitted to a sigmoid equation (Equation 1). (A: Saenuri, B: Seolgaeng, C: Baromi2, D: 22°C, 28°C in Baromi2, E: 18°C in Baromi2).
Table 4.
Parameters of the sigmoid equation relating grain weight of rough rice to the effective accumulated temperatures during the grain filling stage.
| Varieties | Parameters | ||||
| a | b | t0 | vmax | R2 | |
| Saenuri |
23.7 (0.66) |
90.3 (11.80) |
188.1 (10.36) |
0.0656 | 0.9541** |
| Seolgaeng |
22.1 (0.73) |
95.8 (14.43) |
188.5 (12.43) |
0.0577 | 0.9368** |
| Baromi2 |
22.1 (1.50) |
112.0 (34.48) | 176.4 (26.76) |
0.0493 | 0.7929** |
|
Baromi2 (22, 28°C) |
22.1 (0.41) |
84.7 (10.74) |
145.2 (8.32) |
0.0652 | 0.9719** |
|
Baromi2 (18°C) |
29.9 (11.12) |
243.5 (90.06) |
416.3 (198.52) | 0.0307 | 0.9803* |
Vmax로 표현되는 최대 등숙속도(dW/dt)는 새누리 0.0656, 설갱은 0.0577, 바로미2 (저온제외)는 0.0652로 품종간 비슷한 속도를 보였다(Table 4). 반면 저온조건에서의 바로미2는 0.0307로 등숙속도가 매우 느린 것을 확인할 수 있었다. 등숙속도가 최대가 되는 시점(t0)도 역시 품종에 따라 145∼189 내외로 비슷한 값을 보였는데, 바로미2의 저온처리구는 416으로 매우 늦어지는 경향을 보였다.
고 찰
고온조건(평균 28°C)에서는 현미천립중이 분질미, 연질미, 일반품종 순으로 크게 감소하였다(Table 2). 새누리가 고온에서 약 6% 감소한 것은 25°C 이상의 고온에서 약 6%의 천립중 감소를 보인다고 한 기존의 보고(Kim, 1985)와 일치한다. 설갱과 바로미2는 새누리보다 각각 2배, 3배이상으로 크게 감소하였다. 또한 전분의 상대적인 함량(%)은 고온조건에서 큰 감소를 보였고 그 경향은 쌀가루용 품종에서 더욱 두드러졌는데(Table 3), 앞서 언급했듯 현미천립중도 비슷한 경향으로 감소하였으므로 절대적인 전분축적량(g)은 차이가 더욱 클 것으로 생각되었다. 현미천립중(g/천립)과 전분함량(%)을 곱하여 전분축적량(g/천립)을 계산한 결과 고온에서 새누리는 9.3%, 설갱은 14.0% 감소하였으나 바로미2의 경우 30% 이상 감소하며 쌀가루용 품종 중에서도 분질미의 전분 축적이 고온에서 매우 저해되는 것을 볼 수 있었다. 한편 단백질의 상대적인 함량은 온도가 상승할수록 증가하였는데(Table 3), 종실당 단백질 축적량(g/천립)을 계산하면 최대 약 0.1 g의 차이에 그쳤으며 고온에서도 비슷한 값을 보였다. 이와 관련하여 30°C 이상의 고온에서 전분 축적이 저해되는 것과는 달리 단백질 축적은 영향을 받지 않았다는 보고가 있으며(Tashiro & Wardlaw, 1991b; Cooper et al., 2008), 이는 고온에서의 단백질 함량의 증가가 실제로 단백질 축적의 증가보다는 전분축적 감소에 따른 종실중의 감소로 인하여 상대적으로 증가한 것이라고 정리할 수 있다. 따라서 본 연구에서 천립중 감소의 주된 이유는 전분 축적량의 감소라고 생각되며 고온조건에서 분질미의 천립중 감소폭이 컸던 것은 일반 품종과는 다른 분질미의 전분축적관련 유전자의 발현이 고온에서 크게 감소했기 때문이라고 추측된다.
유효적산온도를 이용해 품종간 등숙속도를 비교하였는데(Fig. 1) 바로미2에서 이상치(outlier)를 보였던 저온(18°C)조건의 값을 떼어내어 각각 분석하였고, 저온에서 등숙속도가 현저히 느린 것을 볼 수 있었다. 그러나 앞서 Table 2에서 보았듯이 최종 천립중은 오히려 22°C보다 증가했던 결과를 미루어보아 초반의 등숙속도를 감안하여 등숙기간을 충분히 확보한다면 초반의 부진을 회복할 수 있는 것으로 판단된다. 이처럼 등숙온도, 특히 저온조건에서 등숙속도의 차이가 나는 이유는 등숙 초반 분질배유의 등숙에 관여하는 유전자의 발현 차이에 기인하는 것으로 보이는데, 이는 추후 유전자 수준에서의 추가 연구가 필요하다고 생각된다.
앞서 설명하였던 것처럼 분질미인 바로미2는 등숙기 고온(28°C)에서는 천립중 및 전분함량이 크게 감소하였으며 온도가 낮아질수록 증가하는 경향을 보였다. 따라서 차후 분질미 재배시 등숙기 평균온도가 고온을 피할 수 있도록 이앙시기를 설정하는 것이 중요할 것이라 사료된다. 다만 저온(18°C)에서 등숙초반의 속도가 현저히 느려졌던 것을 감안하였을 때 너무 늦게 이앙할 경우 등숙기간을 충분히 확보하지 못할 수 있으므로 주의해야 될 것이라 판단된다.
적 요
쌀가루용 품종의 등숙기 온도에 따른 등숙특성 변이를 구명한 내용은 다음과 같다.
1. 고온(28°C)에서는 바로미2의 천립중이 21% 감소하면서 새누리(6% 감소)와 설갱(12% 감소)에 비해 크게 줄어들었으며, 전분함량의 감소 또한 바로미2에서 10% 이상으로 새누리와 설갱의 1∼3% 감소와 대비되었다.
2. 저온(18°C)에서는 새누리나 설갱에 비해 바로미2의 등숙 속도가 매우 감소하였으며 특히 등숙 초반에 종실중의 증가가 매우 느렸다. 그러나 최종 천립중 및 전분함량은 22°C보다도 약간 증가하며 다른 품종과도 비슷한 것을 확인하였다.
3. 앞서 언급한 것처럼 쌀가루 가공용 품종 중 분질미(바로미2)는 등숙기 온도에 민감하기 때문에 추후 분질미 재배시 등숙기에 고온을 피할 수 있도록 이앙시기를 늦추는 편이 좋으며, 다만 과도하게 늦출 경우 저온조건에서 등숙기간을 제대로 확보하지 못하여 등숙이 충분히 되지 않을 수 있기 때문에 주의가 필요하다.
