

콩(Glycine max (L.))은 우리나라에서는 중요한 식량작물의 하나로 재배되어 왔으며, 단백질 함량이 30%이상으로 인류 에게 있어 우수한 단백질 공급원이다(Rahman et al. 2011). 하지만 재배면적은 1990년에 152,265 ha에서 2007년에는 76,267 ha로 재배면적이 줄어들고 있으며 자급률도 1990년 에는 20.1%에서 2013년에는 9.7%로 감소하였다(STATISTICS KOREA). 따라서 콩의 자급률 증대를 위해 최근에 쌀 재고 량이 증가하면서 논에서의 콩 재배 면적이 증가하고 있는 추세이다. 논 콩 재배면적은 1990년에 6,575 ha에서 2014년 에는 10,785 ha로 증가하였다(STATISTICS KOREA).
우리나라의 기상 및 재배 환경을 고려하면 콩의 파종기인 5월에는 강우가 부족하고, 콩의 영양생장기와 개화기인 7~8 월에는 장마로 인하여 습해가 일어나기 쉬워 논에서의 콩 재배는 콩 생육기간 중의 습해가 안정생산의 제한적 요인이 될 것이다. 토양이 과습 할 때 식물이 장해를 받는 것은 토 양 중의 산소가 부족하기 때문에 작물의 생리작용이 저해될 뿐만 아니라 유해 물질들이 생기는데 기인한다(Bruce et al., 1992; Lemke-Keyes and Sachs, 1989). Heydecker et al. (1969)는 식물체는 종자 발아 시 각기 다른 토양 내 환경조 건들을 스트레스로서 감지하게 된다고 하였다. Pollock and Ross (1972)는 스트레스의 요인은 유전적인 성질과 종자의 생리조건에 따라서 다르다고 하였으며, 생리적인 스트레스 를 야기하는 인자에는 환경인자(수분, 대기가스, 온도, 광, 이온), 화학적 인자(영양 및 식물독성물질) 및 물리적인 인 자(토양경도 및 토성) 등이 있다고 하였다. 여러 가지 stress 중에서도 논콩 재배시 발아기에 가장 직면하기 쉬운 것은 수분장애이다. 따라서 장애로 인한 생리적 변화는 장애를 극복하기 위한 유전기구를 작용시켜 새로운 단백질을 합성 하거나, 대사과정을 변경시켜 환경장애에 대응한다. 그러나 이러한 대응기작에 대해서는 아직 정확히 밝혀지지 않았다.
본 연구에서는 단백질 발현양상을 비교분석하기 위하여 넓은 등전점영역의 단백질을 높게 분리할 수 있는 능력을 가지고 있는 이차원전기영동 사용하여(Cho et al., 2014) 콩 의 발아전 침종처리가 발아에 미치는 영향을 규명하고 발아 과정에서 발현되는 단백질을 비교분석하고 질량분석기를 이용하여 단백질을 동정함으로써 발아단계에서의 유전적 기작을 이해하기 위한 기초자료를 얻고, 침종 후 발아종자 와 미발아종자간 단백질 발현의 차이를 분석하여 콩 발아단 계에서의 대사적 변화를 이해하기 위한 기초자료를 제공하 고자 하였다.
재료 및 방법
식물재료
침종 일수에 따른 종자의 단백질 발현 양상 비교분석에는 소백나물콩, 태광콩, 장수콩, 황금콩, 단엽콩, 단원콩, Peking 을 사용하였으며 침종에 따른 발아와 미발아에 따른 종자의 단백질 발현 양상 비교분석에는 장수콩, 남해콩, 팔도나물 콩, 신팔달콩 2호, 익산콩, 흑청콩을 사용하였다. 모든 공시 품종들은 충북대학교 부속농장 전작포장에 파종하였으며, 시비는 콩 복비 50 kg/10a (N : P2O5 : K2O = 3 : 3 : 3.4 kg/10a)을 전량기비로 하였으며, 기타 재배관리는 콩 표준 재배법에 준하였다.
단백질추출 및 정량분석
이차원전기영동을 위한 단백질 추출을 위하여 개화 후 시기 별로 채집된 시료의 지방을 제거하기 위하여 85%의 n-hexane 을 사용하였다. 지방을 제거한 시료 0.1 g에 lysis buffer [4.8 g Urea, 0.2 ml NP-40, 0.2 ml Ampholyte pH 3.5-10, 0.5 ml 2-mercaptethanol, 0.5 g PVP-40/10 ml] 1 ml를 첨 가하여 막대사발에서 혼합한 후 15,000 rpm에서 15분간 원 심분리하여 상등액을 취하고, 상등액을 분석에 사용하였다.
단백질 함량 분석은 Bradford (1976)의 방법에 595 nm에서 분광광도계(SHIMADZU, UV-1700)를 사용하여 측정하였다.
단백질 발현 양상비교를 위한 이차원전기영동
이차원전기영동(O'Farrell, 1975)은 넓은 등전점영역의 단 백질을 높게 분리할 수 있는 능력을 가지고 있는 일차원(등 전점) 전기영동 NA-1313 (Nihon-Eido)과 이차원 전기영동 은 NA-1130 (Nihon-Eido)을 사용였다. 일차원전기영동은 등전점전기영동으로 사용된 겔의 조성은 4.8 g Urea, 1.6 ml 30% Acrylamide solution, 2.84 ml DDW, 0.2 ml 10% NP-40, 0.25 ml Ampholyte pH 3.5-10, 0.25 ml Ampholyte pH 5-8, 15 μl 10% APS, 10 μl TEMED이다. 일차원 전기영동이 후 평형화가 끝난 겔을 이차원전기영동을 하였다. 이차원전 기영동은 12% separating gel (9.6 ml DDW, 17.7 ml 30.0% acrylamide solution, 16.4 ml 1.0M Tris-HCl (pH 8.8), 300 μl 10% Ammonium persulfate, 50 μl TEMED)과 5% Stacking gel (2.0 ml DDW, 1.0 ml 30.0% acrylamide solution, 3.0 ml 0.25M Tris-HCl (pH 6.8), 30 μl 10% Ammonium persulfate, 20 μl TEMED)을 사용하였으며 수행하였다.전기영동이 끝 난 후 겔은 은 염색법을 사용하였다. 은 염색의 경우에는 Amersham사의 PlusOne Silver Staining Kit, Protein을 사 용하여 염색을 하였다.
질량분석을 이용한 단백질 동정
이차원전기영동 후 겔에서 원하는 spot을 적당한 크기(1 ×1 mm)로 잘라서 e-tube에 넣었다. 겔을 탈색하기 위해서 30 mM Potassium ferricyanide/100 mM sodium thiosulfate (1:1[V/V]) 100 μl를 첨가하여 흔든 후 원심분리를 실시한 후 상등액을 버렸다. 3차 증류수를 100 μl를 첨가한 후 원 심분리를 하여 다시 상등액을 버렸다. 이 과정을 3번 반복 하였다. 100% ACN 100 μl를 첨가하여 5분 동안 섞은 후 원심분리하여 상등액을 버렸다. 원심분리건조기에서 10분 정도 건조시켰다. Reduction 과정은 10 mM DTT/100mM ABC 100 μl를 첨가하여 56°C에서 1시간 반응을 시켜 원심 분리 후 상등액을 버렸다. 55 mM IAA/100 mM ABC 100 μl를 첨가하여 암조건에서 40분 반응을 시킨 다음 원심분리 후 상등액을 버렸다. 3차 증류수를 IAA의 2~3배를 첨가하 고 섞은 뒤 원심분리하여 상등액을 버렸다. 이 과정을 2번 반복하였다. 100% ACN 100 μl를 첨가하여 5분 동안 섞은 뒤 원심분리하여 상등액을 버렸다. 다시 원심분리건조기에 서 10분간 건조시켰다. 7~8 μl (0.1 μg/μl) trypsin enzyme 를 첨가시켰다. 양이 적을 때에는 50 mM ABC 40 μl를 더 첨가시켰다. 37°C에서 12~17시간 놓아두었다. 반응이 끝난 e-tube는 원심분리 후 상등액을 500 μl tube에 넣고 gel이 든 e-tube에는 50% ACN/5% TFA 50 μl를 첨가하였다. 25°C 실온에서 40분간 놓아두고 원심분리하여 500 μl tube에 넣 었다. 다시 gel이 든 e-tube에 100% ACN 50 μl를 참가하여 원심분리 후 상등액을 500 μl tube에 넣은 후 원심분리건조 기에서 완전히 말린다. CBB 염색의 경우는 30% MeOH 200 μl를 첨가하여 세척하고 50% Acetonitrile/10mA Ammonium bicarbonate 200 μl를 첨가하여 탈색하며 trypsin enzyme을 15 μl를 넣었다. In-gel digestion 후 건조된 시료에 0.5% trifluoroacetic acid (TFA) 10 μl을 가하여 섞은 뒤 원심분 리를 하였다. Zip tip C18 (Millipore, Bedford, MA, USA) 을 wetting solution (50% ACN/0.5% TFA/DW)으로 5번 세척한 후 0.5% TFA/DW로 10번 세척하였다. Pipette을 시 료가 녹아있는 e-tube에 넣고 10번 상하로 통과시켰다. 다 시 0.5% TFA/DW로 10번 세척하였다. 50% ACN/0.5% TFA/matrix/DW 4 μl를 취하여 plate에 1 μl씩 옮긴 후 건조 시켜 단백질 동정을 하였다. 단백질 동정에는 대전 기초과학연 구소의 Voyager-DE STR MALDI-TOF mass spectrometry를 사 용하였으며 instrument type는 MALDI-TOF-TOF로 하였다.
결과 및 고찰
침종 일수에 따른 단백질 발현 양상 비교분석
이차원전기영동을 이용하여 침종 일수에 따른 단백질 발현 양상을 비교분석하였다. 일반적으로 침종 일수에 따라 단백 질 발현 정도가 증가하는 것을 확인 할 수 있었다. 하지만 전 반적인 단백질 발현양상은 침종 일수에 따라 크게 변하지 않 았다. 침종일수에 따라서 단백질 발현정도에 차이를 보이는 단백질 spot들을 선발하고 질량분석을 통하여 동정하였다.
소백나물콩에서는 단백질 발현정도가 침종일수가 증가하 면서 3일째에는 감소하였다가 4일째에는 다시 증가하는 SB-1, SB-2, SB-3이 동정하였다(Fig. 1). 태광콩에서는 새 로 발현하는 단백질 spot T-1과 T-2를(Fig. 2), 장수콩에서 는 단백질 발현정도가 증가하는 J-1과 J-2(Fig. 3)을 동정하 였다. 또한, 황금콩에서도 침종일수에 따라 단백질 발현정 도가 증가하는 H-1, H-2, H-3, H-4 및 H-5를(Fig. 4), 단엽 콩에서는 침종 1, 2일 보다 3, 4일에서 단백질 발현정도가 크게 증가한 DY-1, DY-2, DY-3 및 DY-4(Fig. 5)를 동정하 였다. 단원콩의 경우에는 침종 3, 4일에 나타나는 단백질 spot DW-1, DW-2, DW-3을 동정되었으며(Fig. 6), peking 에서는 침종일수에 따라 단백질 발현정도가 증가하는 spot P-1, P-2, P-3과 침종 4일에 단백질 발현정도가 감소하는 P-4를 동정하였다(Fig. 7).
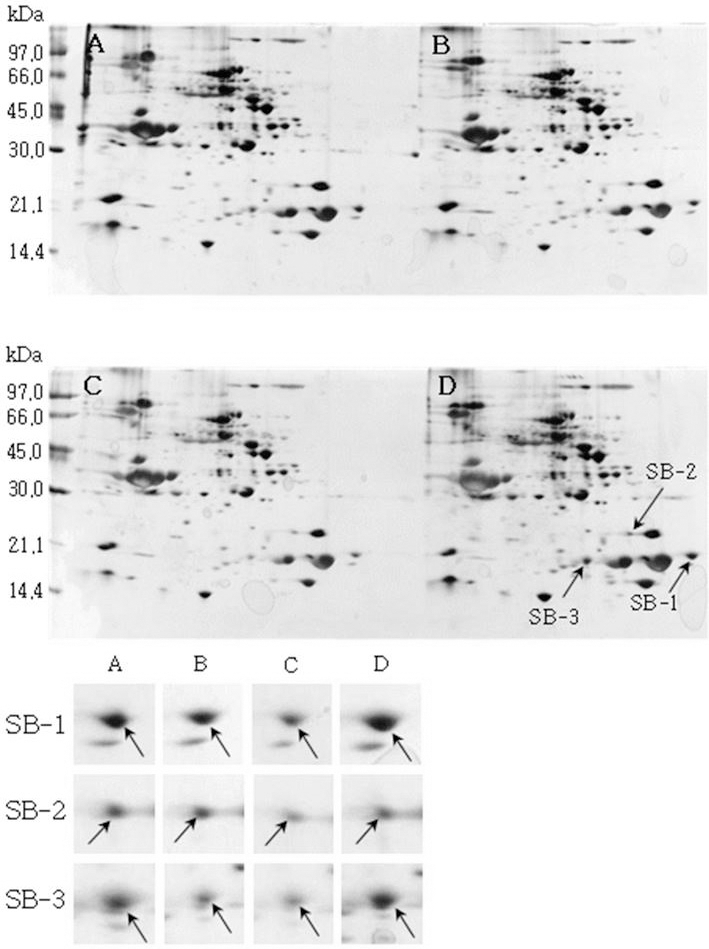
Fig. 1.
Reference 2-DE gel map from Sobaeknamul-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers. (A: 1, B: 2, C: 3, D: 4 days after soaking).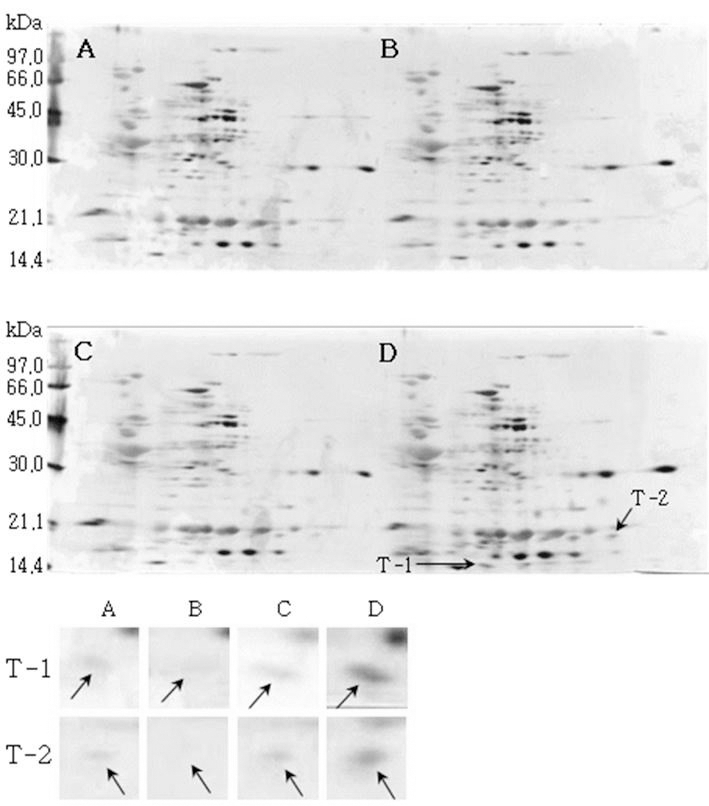
Fig. 2.
Reference 2-DE gel map from Taekwang-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).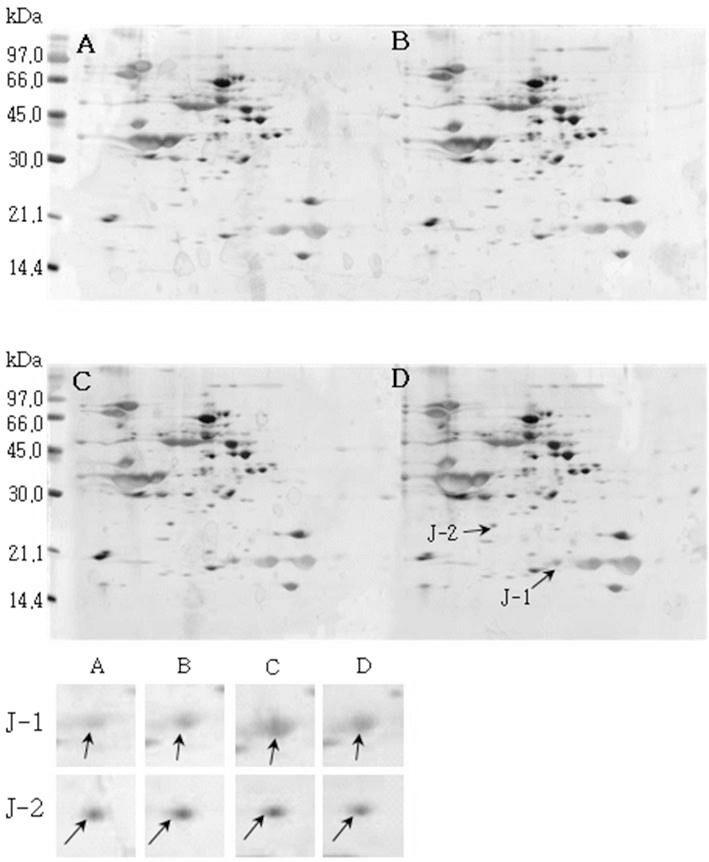
Fig. 3.
Reference 2-DE gel map from Jangsu-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).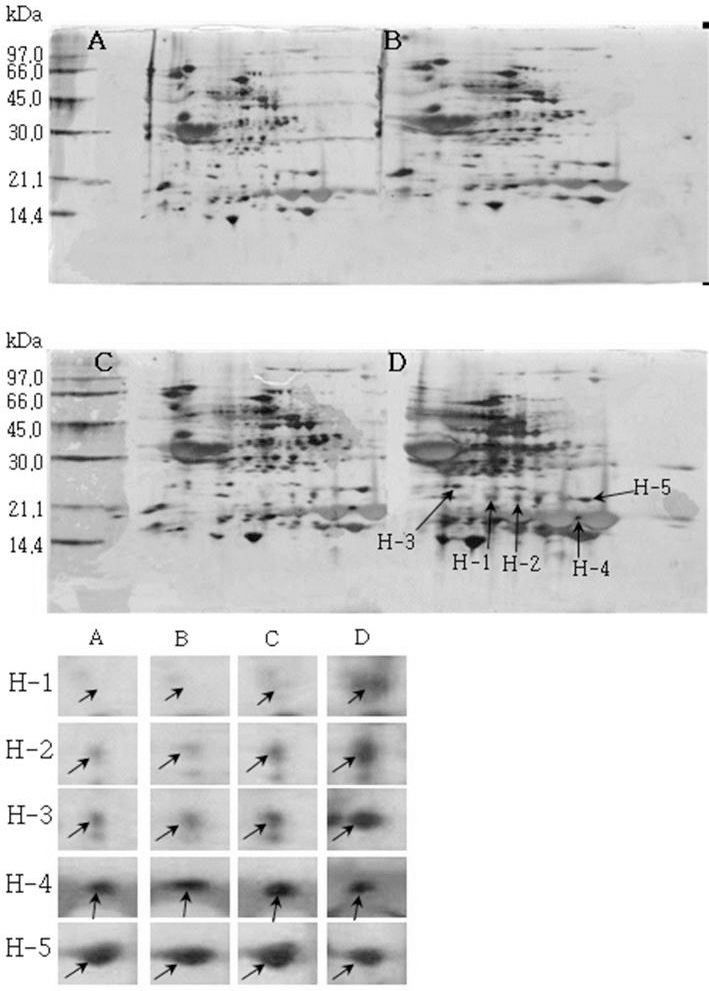
Fig. 4.
Reference 2-DE gel map from Hwangkeum-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).
Fig. 5.
Reference 2-DE gel map from Danyeop-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).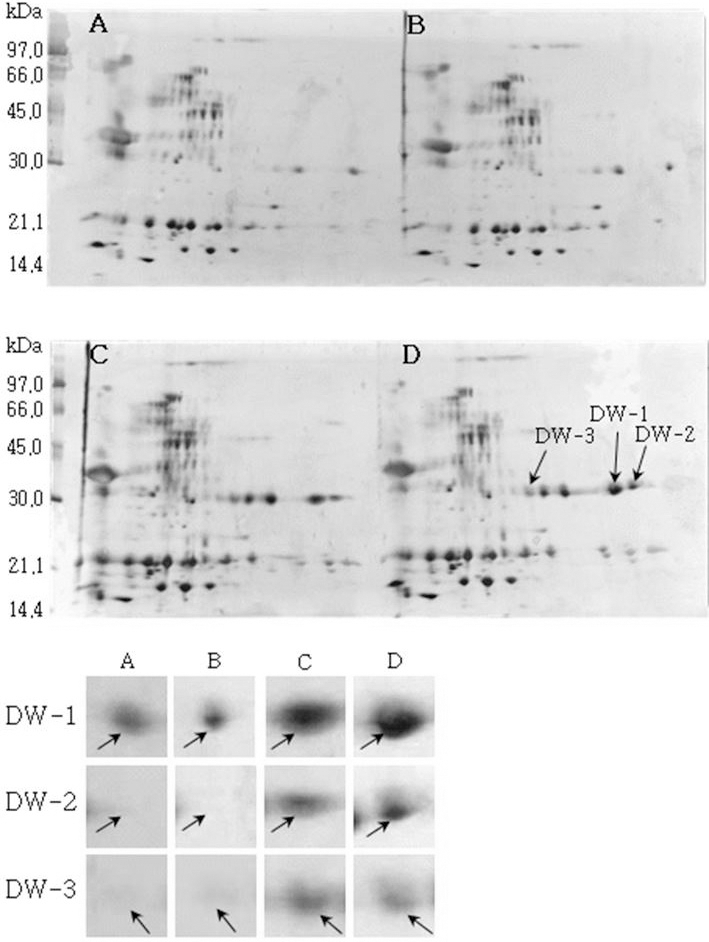
Fig. 6.
Reference 2-DE gel map from Danwon-kong in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).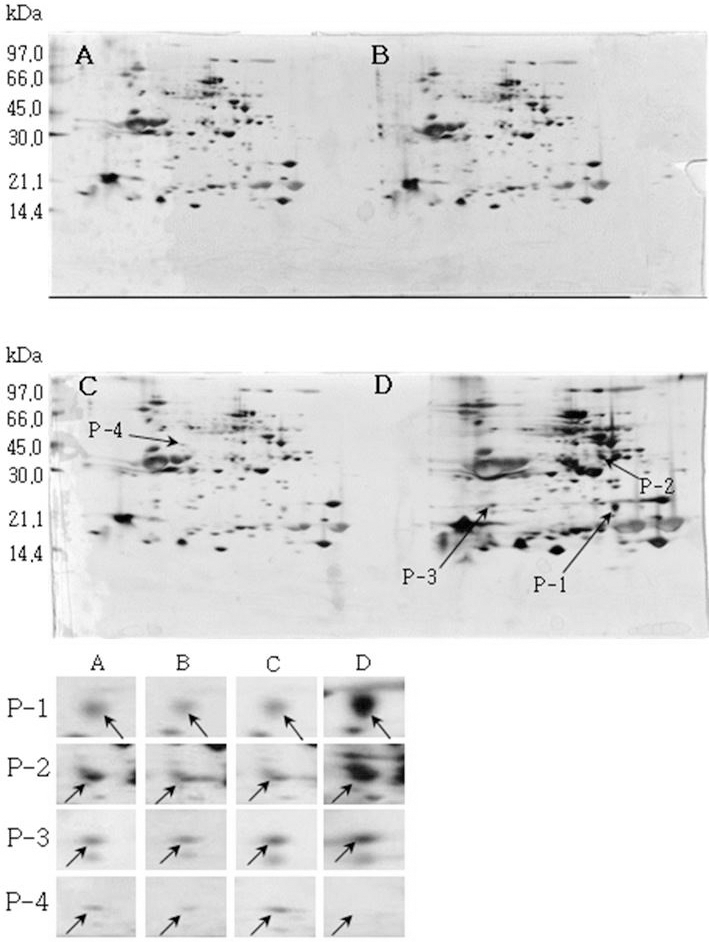
Fig. 7.
Reference 2-DE gel map from Peking in soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: 1, B: 2, C: 3, D: 4 days after soaking).침종일수에 따라서 발현의 차이를 보이는 spot에 대하여 NCBI와 ExPASy에서 동정한 단백질은 Table 1와 같다. 질량분 석을 통해 동정한 단백질 spot 중 SB-1은 Putative transcription elongation factor로, 그리고 SB-3는 Myb oncoprotein homolog {clone 305}로 확인되었다. SB-2, DY-4는 미지의 단백질, 그리고 T-2, J-1, DY-2, DW-1 및 DW-3은 NCBI의 검색결 과 이름이 알려지지 않은 단백질이었지만, Swiss-Prot의 검 색결과 T-2, J-1, DY-2는 Glycinin G1으로, DW-1과 DW-3 는 Basic 7S globulin으로 확인되었다. J-2, H-1, H-2, DY-1, DY-3와 P-1은 Allergen Gly m Bd 28K로 확인되었는데 이 단백질은 콩에 존재하는 주요한 Allergen 단백질 중에 하나 이다(Tsuji et al., 1997). T-1은 MADS box protein 26으로 나타났고(Acton et al., 1997), H-3과 H-4는 Glycinin으로 확인되었다. 특히, H-5는 P24 oleosin isoform B으로 지질 체를 고정시키는 역할을 하며, 발아단계에서 지질의 조절 signal를 조절하는 단백질로 확인되었다(Loer and Herman, 1993). DW-2는 콩의 주요 저장 단백질인 7S seed globulin precursor로 확인되었는데 이 단백질은 높은 분자량과 낮은 분 자량을 포함하는 종자 저장단백질이다. P-2는 Seed maturation protein PM34로 종자 성숙과 관련된 단백질로, P-3은 Lipid transfer protein precursor로 지질수송단백질로 확인되었다 (Kader, 1996). 생육과정에 필요한 아미노산을 공급하기 위 해 저장 단백질이 종자 발아과정에서 소비된다고 하였다 (Krishnan, 2002).
Table 1.
List of identified proteins from 2-DE gel on different soaking daysusing MALDI-TOF-TOF mass spectrometry.
| Spot no. | pI | MW | Protein Name | Genebank Identifiera) | Protein Accession No.b) |
|---|---|---|---|---|---|
| SB-1 | 9.35 | 41188 | Putative transcription elongation factor | gi50919393 | - |
| SB-2 | 9.12 | 22359 | Unknown protein | gi18403720 | - |
| SB-3 | 4.85 | 1393 | Myb oncoprotein homolog{clone 305} | gi256824 | - |
| T-1 | 6.25 | 24888 | MADS box protein 26 | gi4105097 | Q9ZTQ9 |
| T-2 | 6.15 | 55472 | Unnamed protein product/Glycinin G1 (precursor) | gi18165 | P04776 |
| J-1 | 6.15 | 55472 | Unnamed protein product/Glycinin G1 (precursor) | gi18615 | P04776 |
| J-2 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| H-1 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| H-2 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| H-3 | 6.15 | 54208 | Glycinin subunit G3 | gi18639 | P11828 |
| H-4 | 5.78 | 55657 | Glycinin A1aBx | gi225651 | - |
| H-5 | 8.89 | 23487 | P24 oleosin isoform B | gi476216 | P29531 |
| DY-1 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| DY-2 | 6.15 | 55472 | Unnamed protein product/Glycinin G1 (precursor) | gi18615 | P04776 |
| DY-3 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| DY-4 | 9.46 | 21948 | Unknown protein | gi15223241 | - |
| DW-1 | 8.61 | 46262 | Unnamed protein product/ Basic 7S globulin(precursor) | gi18543 | P13917 |
| DW-2 | 8.68 | 46264 | 7S seed globulin precursor | gi1401240 | P13917 |
| DW-3 | 8.61 | 46262 | Unnamed protein product/ Basic 7S globulin(precursor) | gi18543 | P13917 |
| P-1 | 5.73 | 52609 | Allergen Gly m Bd 28K | gi12697782 | Q9AVK8 |
| P-2 | 6.6 | 31747 | Seed maturation protein PM34 | gi96221530 | Q9LLQ6 |
| P-3 | 9.36 | 11798 | Lipid transfer protein precursor | gi13507262 | Q9ATH2 |
| P-4 | 9.21 | 1347 | Hypothetical protein - Soybean | gi7488680 | - |
침종 후 발아와 미발아에 따른 단백질 발현 양상 비교분석
모든 공시 품종들에서 발아한 종자에서 전반적으로 단백 질 발현정도가 발아하지 않은 종자에 비하여 높게 확인되었 으나 소수의 단백질 spot들의 단백질 발현정도는 미발아 종 자에서 높게 확인되었다. 발아한 종자와 발아하지 않은 종 자의 2-DE gel pattern은 Fig. 8, Fig. 9, Fig. 10, Fig. 11, Fig. 12 및 Fig. 13와 같다.
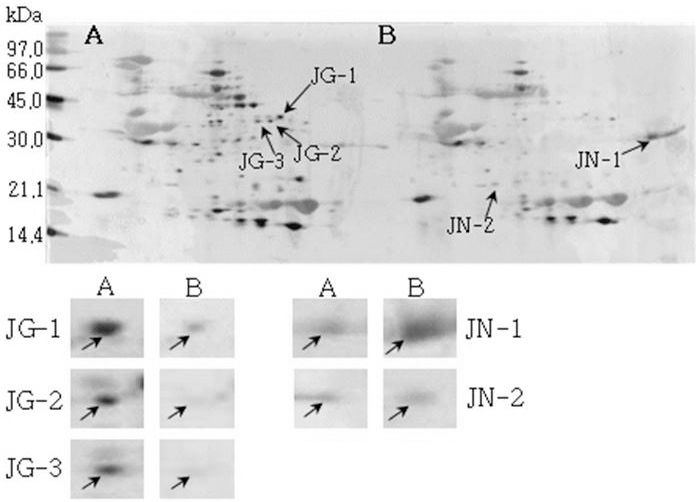
Fig. 8.
Reference 2-DE gel map from Jangsu-kong after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated seeds).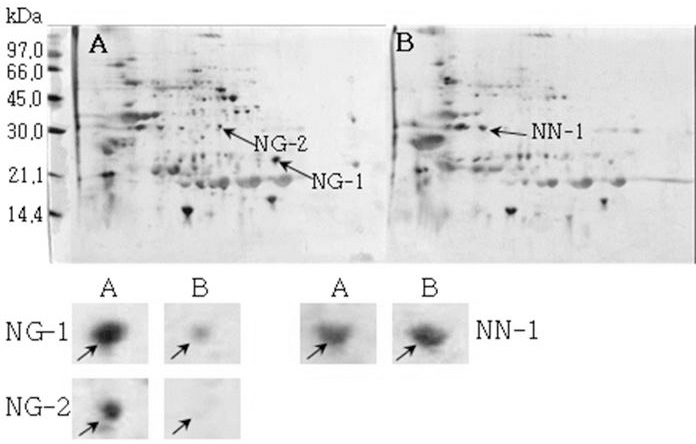
Fig. 9.
Reference 2-DE gel map from Namhae-kong after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated seeds).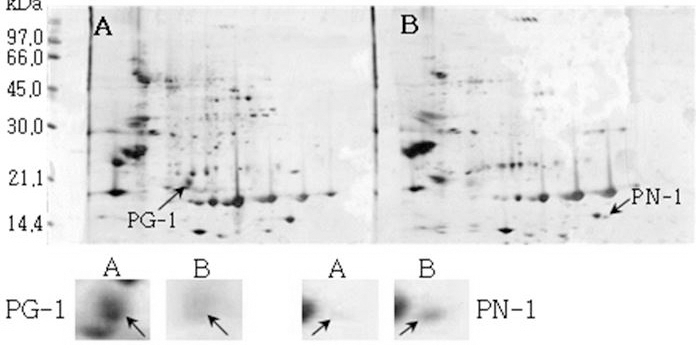
Fig. 10.
Reference 2-DE gel map from Paldonamul-kong after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated seeds).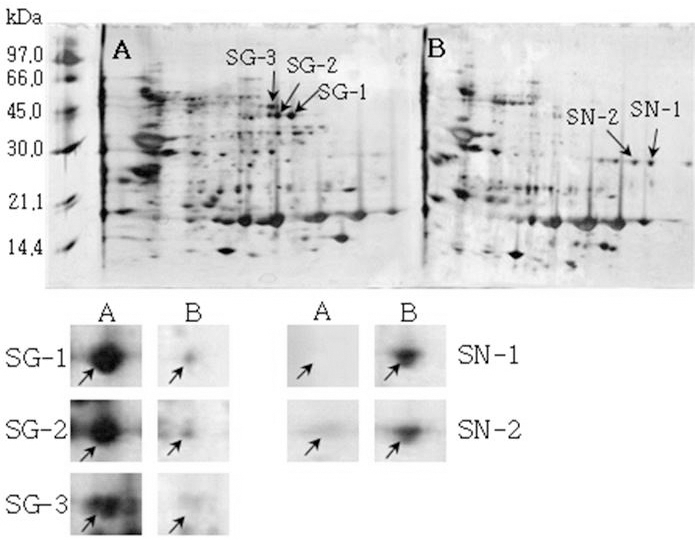
Fig. 11.
Reference 2-DE gel map from Sinpaldal-kong 2 after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated seeds)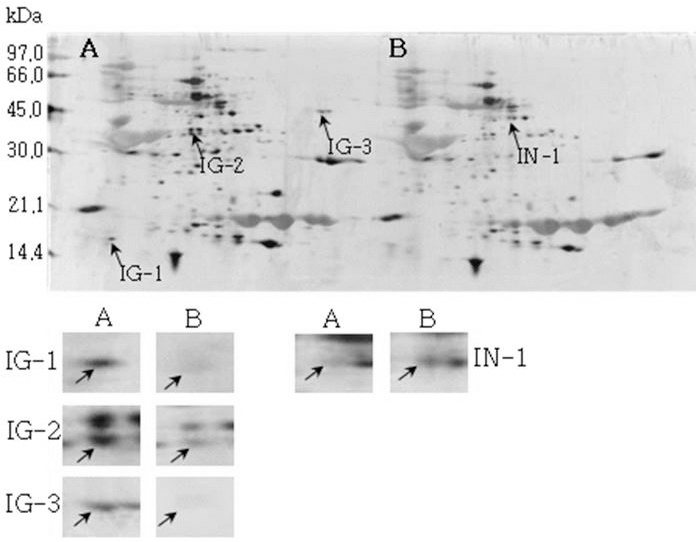
Fig. 12.
Reference 2-DE gel map from Iksan-kong after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated).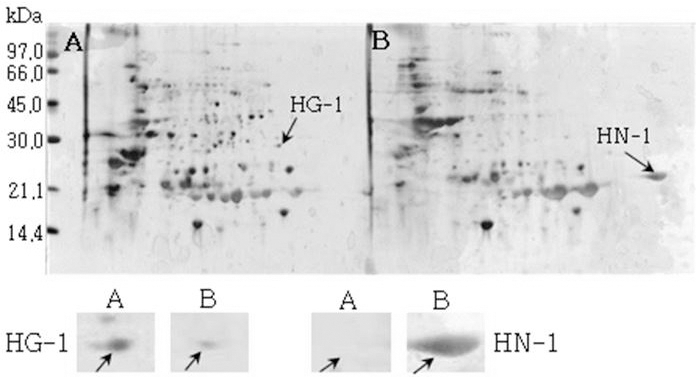
Fig. 13.
Reference 2-DE gel map from Heukcheong-kong after soaking conditions. Proteins were extracted from seeds, separated by 2-DE, and visualized by silver staining. The MW of each protein was determined by 2-DE markers (A: Germinated, B: Un-germinated).발아한 종자와 발아하지 않은 종자의 2-DE gel중에서 발 아한 종자에서만 특이하게 발현한 spot과 발아하지 않은 종 자에서만 특이하게 발현한 spot을 동정할 수 있었다. 장수 콩의 발아한 종자 gel에서는 분자량이 30.0 kDa과 45.0 kDa 사이에 위치한 spot JG-1, JG-2 및 JG-3을 동정하였고, 발 아하지 않은 종자에서는 분자량이 30.0 kDa과 21.2 kDa에 위치한 JN-1과 JN-2를 동정하였다. 남해콩에서는 발아한 종자의 gel에서는 NG-1과 NG-2를 동정하였고, 발아하지 않은 종자에서는 NN-1을 동정하였다. 팔도나물콩의 발아한 종자에서는 PG-1을 그리고 발아하지 않은 종자에서는 PN-1 을, 신팔달콩 2호의 발아한 종자에서는 분자량이 45.0 kDa 주위에 위치한 SG-1, SG-2 및 SG-3과 발아하지 않은 종자 에서는 분자량이 27 kDa에 위치한 SN-1과 SN-2를, 익산콩 의 발아한 종자에서는 IG-1, IG-2 및 IG-3을 그리고 발아하 지 않은 종자에서는 IN-1을, 흑청콩의 발아한 종자에서는 HG-1과 발아하지 않은 종자에서는 HN-1을 동정하였다. 발 아한 종자와 발아하지 않은 종자에서 동정한 단백질중에서 발아한 종자에서 동정된 단백질은 Table 2와 같다. JG-1은 Ubiquitin extension protein으로 단백질 분해과정과 관련된 단백질 확장 단백질로 확인되었고(Callis et al., 1990), JG-2 는 조직 분해효소활성제로 추정되는 Cgi67 serine protease precursor로 확인되었다. JG-3과 IG-1은 NCBI의 검색결과 미지의 단백질이었지만, Swiss-Prot의 검색결과 가상의 단 백질로 확인되었고, NG-1과 IG-3도 가상의 단백질로 확인 되었다. NG-2는 Methllothionein으로 금속과 관련된 단백질 (Coyle et al., 2002), PG-1은 글리신 전구체, SG-1은 NCBI 결과 unnamed 단백질이었지만, Swiss-Prot에서는 Alcohol dehydrogenase 1이라는 단백질로 확인되었는데(Christie et al., 1991), 이는 Crawford and McManmon (1971)의 과습으로 인해 식물에서는 Alcohol dehydrogenase, Malic dehydrogenase, Lactic dehydrogenase, Pyruvate kinase 등 효소들이 발생하여 과습상태를 극복한다고 한 것과 같이 침종으로 인해 Alcohol dehydrogenase 1 단백질이 발생한 것으로 생각된다. SG-2는 효소와 조효소를 조절하는 ATP binding 단백질(Higgins et al., 1986), SG-3은 단백질 분해효소인 inhibitor II, IG-2는 엽록소 환원효소인 엽록체 subunit (Li et al., 1993), 그리고 HG-1은 DNA와 zinc ion을 binding하는 단백질로 각각 확 인되었다(Harrison, 1991; Miller et al., 1985).
Table 2.
List of identified proteins from germinated seeds of soybean using MALDI-TOF-TOF mass spectrometry.
| Spot no. | pI | MW | Protein Name | Genebank Identifiera) | Protein Accession No.b) |
|---|---|---|---|---|---|
| JG-1 | 9.81 | 17676 | Ubiquitin extension protein | gi71913239 | Q45FL8 |
| JG-2 | 8.96 | 42878 | Putative TPA: Cgi67 serine protease precursor | gi34905260 | Q84SE0 |
| JG-3 | 8.24 | 35062 | Unknown protein/Hypothetical protein | gi26449463 | Q8GZ68 |
| NG-1 | 9.64 | 29062 | Hypothetical protein | gi51091686 | Q69L50 |
| NG-2 | 4.11 | 4537 | Methallothionein | gi3342194 | O81528 |
| PG-1 | 5.6 | 57921 | Glycinin precursor | gi121280 | P04347 |
| SG-1 | 6.08 | 41146 | Unnamed protein product/ Alcohol dehydrogenase 1 | gi21951 | P13603 |
| SG-2 | 5.94 | 107304 | ATP binding | gi15217940 | - |
| SG-3 | 4.87 | 8058 | Inhibitor II', proteinase | gi223095 | - |
| IG-1 | 11.52 | 13044 | Unknown protein/Hypothetical protein | gi50909749 | Q6Z5I6 |
| IG-2 | 6.13 | 1474 | Chloroplast subunit of light independent protochlorophyllide reductase | gi18308981 | Q36622 |
| IG-3 | 8.61 | 6791 | Hypothetical protein | gi51091687 | Q69L49 |
| HG-1 | 7.99 | 18417 | DNA binding/zinc ion binding | gi18403408 | - |
발아하지 않은 종자에서 동정된 단백질은 Table 3과 같다. JN-1, JN-2 및 NN-1은 Metallothionein-like protein으로 금 속과 관련된 단백질로 확인 되었고, PN-1과 SN-2는 NCBI 의 검색결과 unnamed 단백질로 나타났는데, Swiss-Prot의 검색결과 Basic 7S globulin으로 확인되었고(Kagawa and Hirano 1989), SN-1은 7S seed globulin precursor, 그리고 IN-1은 지베렐린을 조절하는 단백질로 각각 확인되었다. 익 산콩의 발아하지 않은 종자에서 나타난 것으로 보아 침종으 로 인하여 발아를 촉진시키기 위해 발현된 것으로 사료된 다. HN-1은 글리신 subunit으로 확인되었다.
Table 3.
List of identified proteins from 2-DE gel on on un-germinated seedsusing MALDI-TOF-TOF mass spectrometry.
| Spot no. | pI | MW | Protein Name | Genebank Identifiera) | Protein Accession No.b) |
|---|---|---|---|---|---|
| JN-1 | 5.07 | 6572 | Metallothionein-like protein | gi2465006 | O23938 |
| JN-2 | 4.8 | 7544 | Metallothionein-like protein type 1, putative | gi77556864 | P47988 |
| NN-1 | 4.59 | 7754 | Metallothionein-like protein | gi5853156 | Q9SPE7 |
| PN-1 | 8.61 | 46262 | Unnamed protein product/Basic 7S globulin precursor | gi18543 | P13917 |
| SN-1 | 8.68 | 46264 | 7S seed globulin precursor | gi1401240 | P13917 |
| SN-2 | 8.61 | 46262 | Unnamed protein product/Basic 7S globulin precursor | gi18543 | P13917 |
| IN-1 | 8.99 | 5296 | Gibberellin regulated protein | gi282740631 | Q84XN5 |
| HN-1 | 5.73 | 54208 | Glycinin subunit G3 | gi18639 | P11828 |
적 요
침종 기간과 침종 후 발아의 유무에 따른 단백질 발현을 이차원전기영동을 이용하여 단백질 발현양상을 확인하고 비교 분석한 결과, 침종 기간과 침종 후 발아의 유무에 따른 단백질 발현 양상은 전반적으로 매우 유사하였으며, 주요 단백질의 발현 에는 차이가 없었다. 침종 기간에 따른 품종 별 단백질 발현 양상은 침종 기간이 길어짐에 따라 단백질 발현정도가 점차 증가되는 것을 확인할 수 있었다. 공시품 종들 중 황금콩, 단엽콩, Peking이 다른 품종들에 비하여 침 종 4일후에 단백질 발현정도가 급격히 증가하는 것을 알 수 있었다. 침종 후 발아 유무에 따른 단백질 발현양상은 소수 의 단백질 spot들을 제외하고는 전반적으로 모든 공시 품종 들에서 발아한 종자에서의 단백질 발현정도는 미발아 종자 에서보다 높게 확인되었다.
