

Cultivation season and yield in crops are changed by flowering date. Day length and temperature are reported as the main elements which effect on flowering time. According to effect of day length on flowering, plants can be classified as three types; longer day length, short day length and neutral plant. Longer and shorter day length stimulate flowering in long-day plant and short day plant, respectively. Day length is not effect on flowering in neutral plant.
Rice is a typical short day length plant. In short day length, endogenous genes related with flowering are expressed then protein complex of florigen is translated, finally young panicle is developed in shoot. The growth period of rice is divided as two parts, vegetative growth phase and reproductive stage. After vegetative growth, panicle initiation is started during reproductive stage. Vegetative growth phase is showed difference according to varieties and also greatly effect on growth duration in rice cultivars (Vergara et al., 1985). The vegetative growth phase is also divided as two phases, the basic vegetative phase (BVP) and the photoperiod sensitive phase (PSP). BVP is not affected by light length. However day length is greatly influence on flowering during PSP.
Day length is not changed according to the environment condition. It is just different according to the latitude. However temperature can be changed by environment condition. According to abnormal temperature condition, heading could be appeared in earlier or later (Fitter et al., 2002). It means that ripening condition of rice, yield and quality also could be changed.
According to global warming, average temperature has been increased. Also occurrences of the abnormal climate such as sudden low or high temperature have been increased (Mueller et al., 2012; Dai et al., 2006). In such a condition, rice cultivar with stable heading date regardless of the environment condition can be a good way to ensure the quality and quantity. Effect of light on vegetative growth phase has been investigated well (James et al., 2012; Vittoria et al., 2013; Yoshitake et al., 2014). However the effect of temperature on vegetative growth phase has not been investigated well. Here we tried to investigate the reason of heading delay by low temperature using several Korea varieties.
MATERIALS AND METHODS
Plant materials
Six kinds of Korean varieties (Oryza sativa sp. Japonica) were used as materials (Table 1). Plants of each material were transplanted into a 1/5000-a wagner pot after germination. Plants had been cultivated in the phytotron until heading was appeared. The temperature condition of the phytotron was created as two conditions, 28°C and 22°C. Temperature was 33°C/23°C for the 28°C and 27°C/17°C for the 22°C condition. To eliminate the light effect on heading, light length was fixed as 12 hours. For measuring dry weight of leaf and shoot, each plant was harvested with three replications then dried for 3 days at 70°C. Leaf number and area in the main tiller except a flag leaf were checked when heading were appeared. Leaf area was measured using LI-3100 (LiCOR Inc.).
Table 1
Rice varieties used as materials.
Plant nutrient content
Plants were cut at ground level, washed then dried for three days at 70°C. Dried plants were divided into leaves, stems and each dry weight was measured. The available phosphate content was measured by the vanadate method (NIAST, 2000) and total nitrogen contents of the plants were measured using the Kjeldahl method (Varley, 1996) using an Auto Kjeldahl Foss 2300 (Foss, Denmark).
RNA extraction and gene expression
For RNA extraction, shoot of each material was harvested and frozen by liquid nitrogen, stored at -80°C. Total RNA was isolated from frozen materials by TRI reagent (Invitrogen, USA). cDNA was synthase using RT PCR kit (Takara, Japan). Real-time PCR was performed using Roter-GeneTM 6000 (Corbett Research, Australia). SYBR Green (SYBR Premix, Takara, Japan) was used as the fluorescent dye according to the instructions. The primer sequence was in Table 2 (Araki, 2006).
Table 2
PCR primer sets for real-time PCR.
RESULTS
Change of heading date under different temperature conditions
The heading date of each variety was checked then the accumulated temperature from sowing to heading was analyzed (Fig. 1). In the 28°C condition, heading was appeared 44~54 days after sowing. In the 22°C condition, heading was delayed compare to the 28°C condition in all varieties. Accumulate temperature until heading (ATH) in the 28°C condition was different according to varieties from 1232°C to 1512°C. Keumo, Chenga and Jungsaenggold were showed a high ATH around 1500°C in the 28°C condition. The ATH of Milyang 255, Younghojinmi, Joun and Hopum were around 1300°C in such a condition. The ATH was increased in the 22°C compare to the 28°C condition. The ATH of Keumo, Cheonga and Milyang 255 were increased about 20% in the 22°C compare to the 28°C condition. The ATH of Jungsaenggold in the 22°C condition was slightly increased about 7% compare to the 28°C condition.
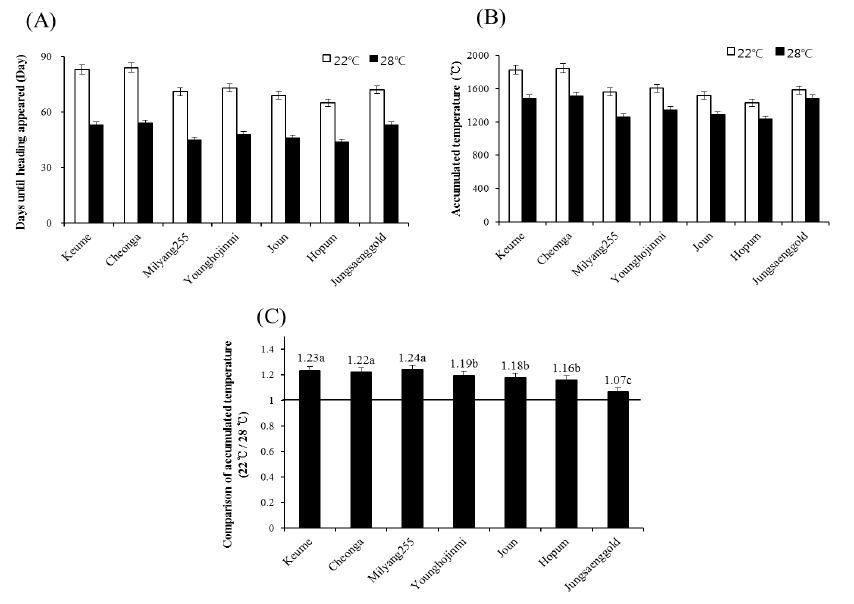
Fig. 1
Change of vegetative growth phase (A), accumulated temperature during vegetative growth phase (B) and comparison of accumulated temperature during vegetative growth phase (C) under different temperature. VGP is considered as period from sowing to heading. Heading date was checked when first heading was appeared. Error bars indicate standard deviations.Plant growth under different temperature conditions
Leaf and shoot dry weights in each material were measured when heading were appeared (Fig. 2). In all varieties, the leaf dry weight were decreased in the 22°C compare to the 28°C condition. The leaf dry weight of Keumo, Cheonga and Joun were showed dramatically reduction in the 22°C compare to the 28°C condition. However, those of milyang 255, Hopum and Jungsaenggold were showed lower decrease compare to other varieties. Shoot dry weights were significantly decreased in Keumo, Cheonga and Jungsaenggold in the 22°C condition. Those of Joun and Hopum were slightly decreased and those of milyang 255, Younghojimmi were slightly increased in the 22°C condition.
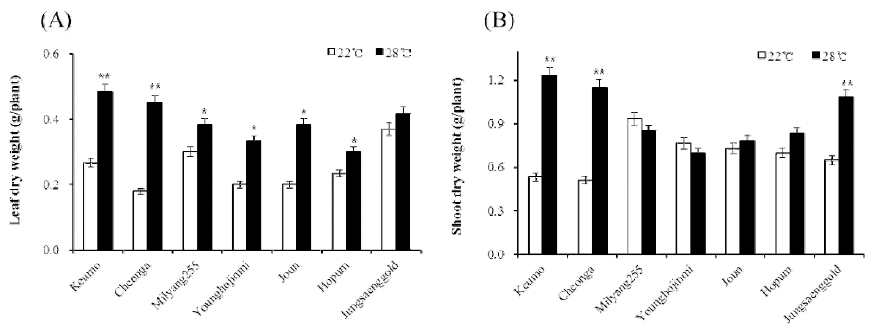
Fig. 2
Change of leaf (A) and shoot (B) dry weight under different temperature conditions. Leaf and shoot day weight was measured when heading was appeared in each condition. Leaf weight was measured in main tiller except a flag leaf. Error bars indicate standard deviations. Significant differences between treatments are indicated with asterisks (**P < 0.01; *P < 0.05).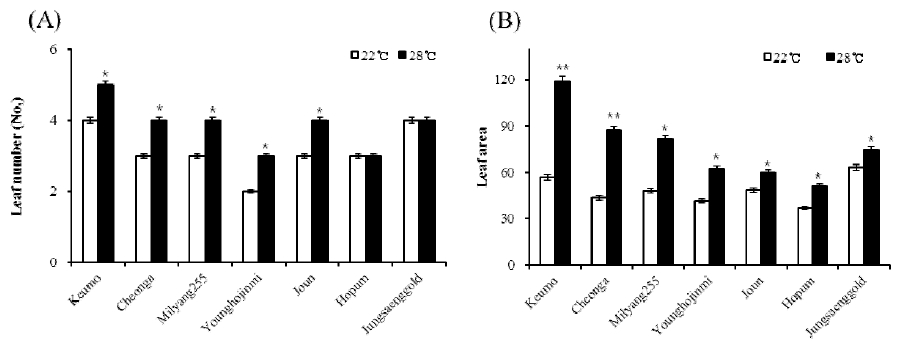
Fig. 3
Leaf number (A) and leaf area (B) of each material under different temperature conditions. Leaf number and area except flag leaf was checked in a main tiller when heading was appeared in each condition. Error bars indicate standard deviations. Significant differences between treatments are indicated with asterisks (**P < 0.01; *P < 0.05).The number and area of leaf in all varieties were decreased in the 22°C condition. Except Hopum and Junsaenggold, leaf numbers were decreased in the 22°C compare to the 28°C condition. The leaf area was showed the biggest decrease in Keumo as the 50% reduction. Those of Cheonga, Milyang 255 and Younghojinmi were showed about the 30 to 40% of reduction. In Joun, Hopum and Jungsaenggold, the reduction of leaf area was the 19, 27 and 15%, respectively.
Nutrient contents in plant under different temperature conditions
For measuring nutrient content in plant, three varieties showing different heading date and leaf growth under different temperature were selected. Nutrient contents such as nitrogen, available phosphate and potassium content were measured when heading was appeared in each condition (Fig. 4). Nitrogen contents (%) in leaf and shoot were higher in the 28°C condition than in the 22°C condition. The leaf nitrogen content of Keumo, Cheonga and Jungsanggold in the 22°C condition were decreased about 20, 25 and 15% compare to the 28°C condition, respectively. In shoot, nitrogen content also decreased in the 22°C compare to the 28°C condition. In the 22°C condition, shoot nitrogen content of Keumo, cheonga and Jungsanggold were decreased about 15, 30 and 10% compare to the 28°C condition, respectively. Available phosphate content was showed different pattern in the leaf and shoot under different temperature conditions. In the 22°C condition, available phosphate content was decreased in leaves, but increased in shoots compare to the 28°C condition. In the 22°C condition, potassium content in leaves was decreased, but potassium content in shoots did not show significant difference compare to the 28°C condition.
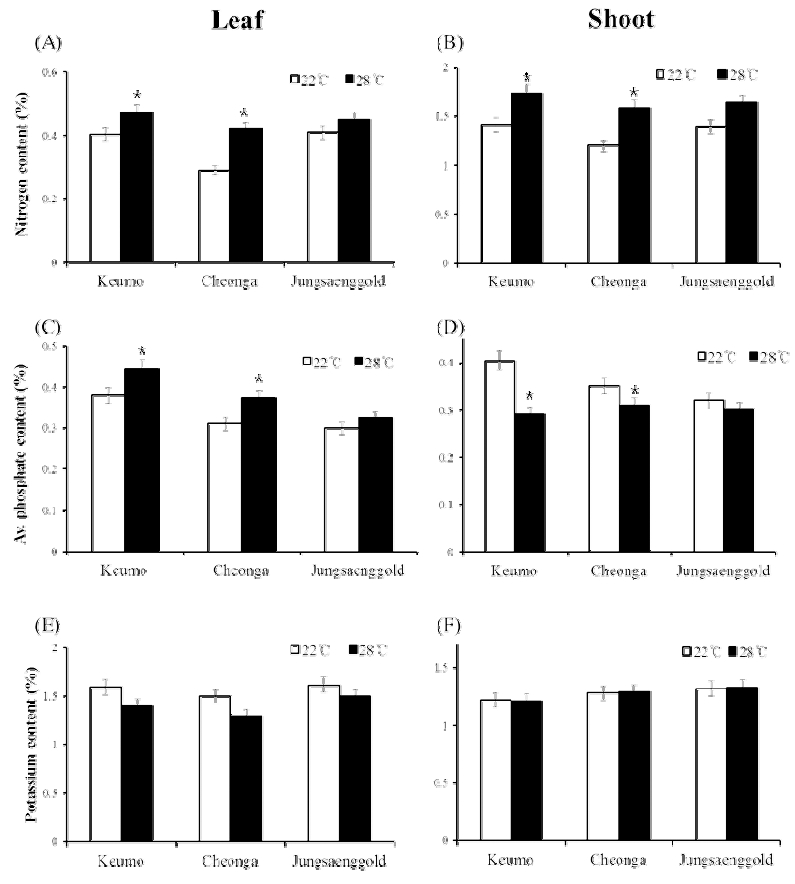
Fig. 4
Nitrogen content (%) in leaf (A) and shoot (B), available phosphate content (%) in leaf (C) and shoot (D), potassium content (%) in leaf (E) and shoot (F) under different temperature conditions. Error bars indicate standard deviations. Significant differences between treatments are indicated with asterisks (**P < 0.01; *P < 0.05).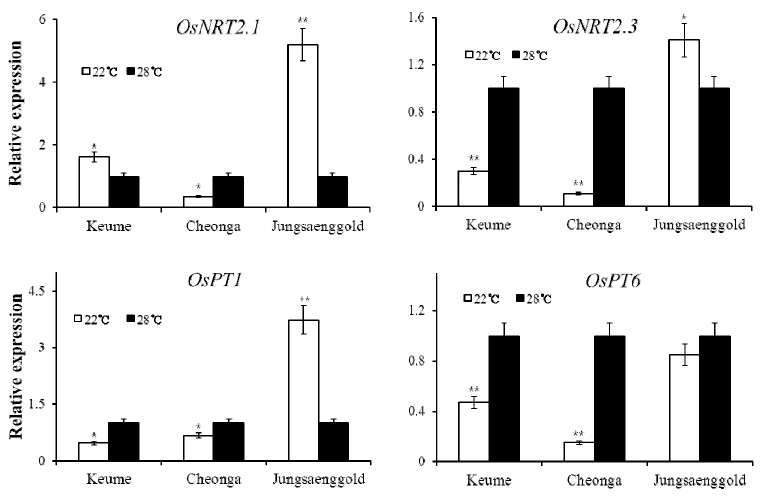
Fig. 5
Change of gene expression in the shoot under different temperature condition. The expression of two kinds of nitrate transporter, OsNRT2.1 and OsNRT2.3, and two kinds of phosphate transporter, OsPT1 and OsPT6, were checked to nutrient uptake ability. Error bars indicate standard deviations. Significant differences between treatments are indicated with asterisks (**P < 0.01; *P < 0.05).Gene expression under different temperature conditions
The expressions of genes related with uptake of nitrogen and available phosphate were checked in the shoot under different temperature conditions. The expressions of OsNRT2.1 and OsNRT2.3 which are nitrate transporter were changed under different temperature conditions. In the 22°C condition, the expression of OsNRT2.1 was increased about 50% in Keumo, 70% decreased in Cheonga compare to 28°C condition. However, OsNRT2.1 was 4 times over expressed in Jungsaenggold under the 22°C condition compare to the 28°C condition. The expression of OsNRT2.3 under the 22°C compare to the 28°C condition was decreased about 70% and 85% in Keumo and Chenga, respectively. However, the expression of OsNRT2.3 in Jungsaenggold was increased about 50% under the 22°C condition compare to the 28°C condition. Both OsPT1 and OsPT6 which are phosphate transporter were down regulated in Keumo and Cheonga under the 22°C condition compare to the 28°C condition. However, OsPT1 in Junsaenggold was overexpressed about 3.2 times more under the 22°C condition compare to the 28°C condition. In the 22°C condition, the expression of OsPT6 in Jungsaengold was decreased about 10 % compare to the 28°C condition.
DISCUSSION
Heading date is important factor of yield and quality in crops. Day length and temperature are reported as one of the main elements which are influence on heading. Compare to day length, temperature effect on heading during growth period has not been investigated well in rice. We cultivated six kinds of Korean rice varieties (Table 1) under high (28°C) and low (22°C) temperature condition in short day length to investigate temperature effect on heading during vegetative growth phase (VGP) (Fig. 1). In the high temperature condition, heading was appeared 45 to 53 days after sowing. In the low temperature condition, VGP of Junsaenggold, Keumo and Cheonga was 53, 53 and 54 days, respectively. However in the low temperature condition, VGP was showed significant difference among varieties. The VGP of Junsaenggold was 72 days in low temperature. The VGP of Keumo and Cheonga were about 83, 84 days in the low temperature condition, respectively.
Analyzing plant growth under different temperature conditions (Fig. 2), the leaf dry weight was generally decreased in the low temperature condition compare to the high temperature condition. However the shoot dry weight was showed a similar pattern among varieties. With those data, we considered that the leaf growth was more strongly related with heading than the shoot growth. We checked change of leaf number and area in a main tiller except a flag leaf under different temperature conditions (Fig. 3). Among varieties, Keumo and Cheonga which were showed significant heading delay under the low temperature condition were also showed significant reduction in the leaf area in such a condition. However the leaf area of Jungsaenggold was decreased about 15% and the leaf area was still wide in the low temperature condition compare to other varieties. Importance of the leaf in flowering has been reported. Song et al. reported that regulated FLOWERING LOCUS (FT) gene in leaves effect on flowering (Song et al., 2013). Corbessier et al. (2007) also reported that FT protein which is the major component of florigen is synthesized in the leaf vasculature then moves to a shoot apical meristem for flowering. We also considered that the reduction of leaf growth in Keumo and Cheonga could be the one of main factors in heading delay under the low temperature condition. On the other hand, more vigorous growth of leaf in Jungsaenggold under the low temperature condition could inhibit severe heading delay. Similarly, Dennett et al. (1978) reported that temperature is effect on the relative leaf growth rate in crops. Thiagarajah et al. (1982) also reported that leaf appearance is increased at high temperature, thus leaf area is decreased at low temperature in corns. In rice, the period of leaf appearance is also reduce under high temperature condition (Ku et al., 2013)
To check the reason of leaf growth difference among three varieties, we investigated the nutrient conditions of plant under different temperature conditions (Fig. 4). Shimono et al. (2012) reported nitrogen uptake is decreased in low water temperature especially during vegetative growth stage in rice. Similarly with Shimono’ report, nitrogen contents in leaf and shoot are decreased under low temperature in all varieties. However the leaf nitrogen content in Jungsaenggold did not show significant difference and the shoot nitrogen content in Jungsaenggold was less decreased under low temperature than other varieties. Available phosphate content in the leaf was decreased in low temperature. However the available phosphate content of leaf in Jungsaenggold were showed less difference between the low and high temperature conditions than other varieties. In a shoot, available phosphate content was increased under low temperature. We considered that high phosphate content in the shoot was due to reduction of the leaf phosphate content. Available phosphate content of the shoot in Junsaenggold was less increased than other varieties. With these data, decrease of nitrogen and available phosphate contents in the leaf under low temperature could be the one of main reasons in reduction of leaf growth. We investigated the expressions of nitrogen and phosphate transporters to confirm the pattern of nutrient uptake under different temperature conditions (Fig. 6). Even nutrients are vigorously uptake by root, nitrogen and phosphate transporter are also showed up regulated patterns in a shoot under the low nutrient condition (Ai et al., 2009; Seo et al., 2008; Fan et al., 2016; Zhenlong et al., 2009). Since plant was cultivated in a pot, we checked genes expression in a shoot to reduce RNA degradation during processing of plant materials. Nitrogen transporter, OsNRT2.1 and OsNRT2.3 were down regulated or showed similar expression under the low temperature condition compare to the high temperature condition in Keumo and Cheonga. However OsNRT2.1 and OsNRT2.3 in Junsaenggold were showed higher expression under the low temperature condition compare to the high temperature condition. Available phosphate transporter, OsPT1 and OsPT6, were showed lower expression in Keumo and Cheonga under the low temperature than the high temperature condition. Even though OsPT6 was slightly down regulated, the expression of OsPT6 in Junsaenggold was still higher than other varieties under the low temperature condition. Over-expressions of OsNRTs and OsPTs have been reported to increase nutrient content in plant (Feng et al., 2011; Osmário et al., 2015; Seo et al., 2008; Wang et al., 2012). Phosphate transporter is generally up regulated to increase phosphate uptake. Among phosphate transporters, OsPT6 was reported to have a role in translocation of phosphate in plant (Ai et al., 2009). The high content of phosphate in shoot of Keumo, Cheonga was considered as the results of reduction in phosphate translocation. With these data, we expected that decrease of nitrogen and available phosphate uptake and translocation under the low temperature condition would reduce leaf growth then leaf growth reduction might cause heading delay. In further study, we will try to investigate the reason of heading delay under low temperature with more definitive study after finding a good genetic resource which shows similar growth pattern regardless of temperature condition.
