

Potato (Solanum tuberosum L.) is the fourth largest staple crop in the world after rice, wheat, and maize. Potatoes are propagated vegetatively, which allows for passage of viral diseases from one generation to the next, making it possible for an entire clonal population to become infected with the same pathogen. Viral diseases are a major constraint to high yield and excellent potato quality. Potato virus Y (PVY) and Potato leaf roll virus (PLRV) are among the most devastating viruses and are spread in most growing areas. Thus, virus-free stocks of plant materials are the basis for the productivity and nutrient value of potatoes (Waterworth and Hadidi 1998). Meristem culture, thermotherapy, and thermotherapy followed by meristem culture are generally the most often used methods applied to obtain virus-free potato plants (Slack and Singh 1998; Faccioli 2001).
Cryopreservation of clonal plant germplasms is becoming a common space-saving technique for long-term and contaminationfree storage of genetic resources (Benson 2008; Wang and Valkonen 2008; Wang et al. 2009). Brison et al. (1997) demonstrated for the first time that cryopreservation can not only be used to conserve germplasm but also to eradicate viruses using the rootstock of Prunus infected with Plum Pox Potyvirus. Since then, cryogenic techniques have been successfully applied to a large number of plant species including agricultural and horticultural crops from temperate and tropical regions (Reed 2002; Panis and Lambardi 2006; Gonzalez-Arnao et al. 2008). Moreover, cryotherapy involving vitrification of shoot tips does not require special equipment in addition to that used in a typical plant tissue culture laboratory. Therefore, we postulate that cryotherapy has the potential to replace more traditional methods such as meristem culture. At the same time, we have succeeded in cryopreserving potato shoot tips grown in vitro (Kim et al. 2006; Yi et al. 2012).
In this study, we evaluated the rates of shoot survival, regrowth, and virus elimination from shoot tip culture and two types of cryogenic procedures, such as droplet-vitrification and encapsulation-vitrification, to eradicate PVY and PLRV from potato shoot tips grown in vitro. We also investigated the effects of shoot tip size on rates of shoot survival, regrowth, and eradication of PVY and PLRV by cryopreservation followed by shoot tip culture.
Materials and Methods
Plant material and shoot tip culture
PVY or PLRV mechanically infected Solanum tuberosum cv. Jasim plants were used. The potato tubers infected with virus were obtained from the Highland Agriculture Research Center, a branch institute of the National Institute of Crop Science, RDA, Pyeongchang, Korea. Stems were taken from potato tubers diagnosed with PVY or PLRV, surface disinfected using 1% hypochlorite solution for 10 min, and washed three times in sterile distilled water. Shoot tips (1.0~1.5 mm) were excised and used as explants to establish a stock culture in vitro in Murashige and Skoog (MS) basal medium (Murashige and Skoog 1962) containing 30 g·L-1 sucrose and 2.2 g·L-1 Phytagel (Sigma-Aldrich, St. Louis, MO, USA) with 0.1 mg·L-1 GA3 and 0.1 mg·L-1 kinetin, followed by incubation at 23 ± 1°C under a 16 h light/8 h dark photoperiod.
Droplet-vitrification cryogenic procedure
A droplet-vitrification procedure was used to freeze the shoot tips. Isolated shoot tips were pre-cultured in liquid MS medium containing 0.3 M sucrose for 7 h, and the shoot tips were further pre-cultured in liquid MS medium containing 0.7 M sucrose for 17 h under the same conditions. The pre-cultured shoot tips were dehydrated in modified plant vitrification solution 2 (PVS2) [37.5% glycerol + 15% dimethyl sulfoxide (DMSO) + 15.0% ethylene glycol + 22.5% sucrose] for 30 min (Table 1). Five drops of dehydration solution were placed on an aluminum foil strip (8 × 20 mm) using a micropipette. One shoot tip was placed in each of the five dehydration solution drops, and the foil strip was immediately plunged into liquid nitrogen. One hour after plunging into liquid nitrogen, two foil strips were transferred to a single 2-mL cryovial that had been filled with liquid nitrogen and stored in a liquid nitrogen tank for more than 1 day. The foil strips were taken out of the cryovials and immediately plunged into a preheated (40°C) unloading solution of 0.8 M sucrose; another 5 mL of room-temperature unloading solution was added after 30 s. The shoot tips were incubated at room temperature for 30 min to facilitate unloading. The thawed shoot tips were washed with 1.0 M sucrose for 30 min at room temperature and postcultured on MS medium supplemented with 0.1 mg·L-1 GA3 and 0.1 mg·L-1 kinetin.
Table 1.
Comparison of the key steps in the droplet vitrification vs. the encapsulation-vitrification protocols for cryopreserving potato shoot tips in vitro.
| Steps | Droplet-vitrification | Encapsulation-vitrification |
|---|---|---|
| Pre-culture | 0.3M(7h)+0.7M(17h) sucrose | 0.25M(24h)+0.5M(24h)+0.75Msucrose(24h) |
| Loading | - | 0.4M sucrose+2M glycerol (90min) |
| Dehydration | Modified PVS2z (30min) | PVS2y (4h) |
| Thawing and unloading | 40°C (30sec)+25°C(30min) in 0.8M sucrose | 40°C (3min)+ 25°C(30min) in 0.8M sucrose |
| Post-culture | MS+GA3+kinetin (0.1+0.1mg·L-1) | MS+GA3+kinetin (0.1+0.1mg·L-1) |
Encapsulation-vitrification cryogenic procedure
Each bead containing one shoot tip was stepwise precultured in increasing sucrose concentrations of 0.25, 0.5, and 0.75 M for 1 day in each concentration. Pre-cultured beads were loaded with a loading solution composed of MS supplemented with 0.4 M sucrose and 2 M glycerol for 90 min at room temperature, followed by dehydration with PVS2 (30% glycerol + 15% DMSO + 15.0% ethylene glycol + 13.7% sucrose; Sakai et al. 1990) for 4 h (Table 1). After dehydration, 10 beads were transferred to a 1.8-mL cryotube and immersed directly in liquid nitrogen for 1 h. Cryotubes that had been preserved in liquid nitrogen were thawed rapidly at 40°C for 3 min. The thawed beads were washed with 1.0 M sucrose for 30 min at room temperature and post-cultured on MS medium supplemented with 0.1 mg·L-1 GA3 and 0.1 mg·L-1 kinetin.
Survival and regeneration assessment
The shoot tips were post-cultured in recovery medium at 23 ± 1°C under dim light for 7 days and then transferred to standard culture conditions to assess viability. Survival rates were evaluated 14 days after cryopreservation by counting the number of shoot tips that were green and swollen (>3 mm). Regeneration rates were estimated 5-6 weeks after cryopreservation by counting the number of shoot tips that had differentiated. Survival and regeneration percentages were averaged for four cultivated potatoes.
Virus detection by reverse transcription-polymerase chain reaction (RT- PCR)
Viruses were tested at three different stages: before shoot tip culture, after shoot tip culture, and after cryogenic treatments. Total RNA was extracted from leaves using the Plant Pathogenic Viral RNA Extraction Kit (Hana Science Co., Seoul, Korea), according to the manufacturer’s instructions. RT-PCR was performed in one reaction tube using the TOPscriptTM One Step RT-PCR kit (Enzynomics, Seoul, Korea). The RT-PCR reaction was done in a 20 μL reaction volume containing 1 μL of 10 pM each primer, 10 μL of the Enzyme mixture (kit), 2 μL extracted RNA, and 6 μL of RNAase-free water. The forward primer 5′-ATGACACAATTGATGCAGGAGGAA-3’ and the reverse primer 5’-GGTGGTGTGCCTCTCTGTGTTCT- 3′ were used for PVY and expected to amplify a 759-nucleotide (nt)-long fragment of PVY-CP. The forward primer 5′- TGGTGTACAACAACCAAGAA-3′ and the reverse primer 5′-CCTTCGTAATTTGGAACTTG-3′ were used for PLRV to amplify a product of 431 nt. RT-PCR amplification was carried out in a thermal cycler (Genepro; Bioer Technology, Tokyo, Japan) using the following program for PVY: cDNA synthesis step (RT reaction) at 50°C for 30 min and initial denaturation step at 95°C for 10 min, 35 cycles at 95°C for 30s, 58°C for 30s, and 72°C for 1 min, followed by a final extension at 72°C for 5 min.
The PLRV program was the following: a cDNA synthesis step (RT reaction) at 50°C for 30 min and initial denaturation step at 95°C for 10 min, 35 cycles at 95°C for 30s, 50°C for 30s, and 72°C for 1 min, followed by a final extension at 72°C for 5 min. The RT-PCR results were analyzed using an automated electrophoresis system (Multina; Shimadzu, Otsu, Japan).
Data analysis and statistical procedures
The results were obtained as average percentages with standard deviations. Each experiment consisted of three replicates per treatment, and each cryovial held 10 samples. The results were analyzed by analysis of variance, and the means were separated using Duncan’s multiple-range test (p < 0.05).
Results and Discussion
Effects of cryogenic methods on shoot recovery and eradication of virus rates
As shown in Table 2, much higher regeneration rates (99.1%) were observed from the shoot tip culture than those of the droplet-vitrification (65.3%) and encapsulation-vitrification (70.5%) procedures. Both cryogenic procedures produced similar regeneration rates. The green-colored cryo-treated shoot tips grew well after 35 days of culture. When axillary buds numbered 3-4 on each explant, they were tested for PVY and PLRV by RT-PCR. The lowest virus eradication rates were observed in the shoot tip culture compared with the droplet-vitrification and encapsulation-vitrification methods. We observed the lowest PVY eradication rate (42.3%) in the shoot tip culture compared with that in droplet-vitrification (91.3%) and encapsulation-vitrification (97.9%). The PLRV results were very similar, as the lowest eradication rate (48.6%) was found in the shoot tip culture compared with that in droplet-vitrification (98.4%) and encapsulation-vitrification (99.7%; Table 2). Fig. 1 and 2 show the health status of ‘Jasim’ potato plantlets obtained from shoot tip culture and cryopreservation followed by shoot tip culture. Six of 10 bands from the shoot tip culture confirmed infection with PVY and PLRV, whereas all plantlets from the cryopreservation followed by shoot tip culture treatments were free of PVY and PLRV.

Fig. 1.
Detecting Potato virus Y (PVY) by reverse transcription–polymerase chain reaction in virus-infected potato stock plants (Jasim) and plantlets regenerated following shoot tip culture and cryotherapy (encapsulation-vitrification). Nucleotides were the expected amplification products from PVY. (M) 100-bp DNA ladder marker; (lanes 1~10) detecting PVY before shoot tip culture; (lanes 11~20) detecting PVY after shoot tip culture; (lanes 21~30) after cryogenic treatment (encapsulation-vitrification).Table 2.
Effects of two kinds of cryogenic treatments on shoot-tip culture survival, regeneration, and eliminating viruses in Solanum tuberosum cv. Jasim.
| Method | Survival (%) | Regeneration (%) | Virus elimination (% virus-free plants) | ||
|---|---|---|---|---|---|
| PVY | PLRV | ||||
| Shoot tip culture | 100 | 99.1±5.3a | 42.3±2.3b | 48.6±3.1b | |
| Cryotherapy | Droplet-vitrification | 99.3 | 65,3±3.1b | 91.3±2.1a | 98.4±2.7a |
| Encapsulation-vitrification | 98.3 | 70.5±4.8b | 97.9±1.5a | 99.7±1.3a | |
Apical meristems of systemically infected plants can typically be rendered virus-free following heat and chemical therapies, after which immunological and molecular diagnostics confirm their disease-free status (van der Want and Diijkstra 2006). Conventional virus elimination methods involve heat treatment (thermotherapy; Kassanis 1950) that can be applied to shoots introduced through tissue culture before they are subjected to meristem culture, which suppresses pathogen titers and enhances virus eradication (Walkey and Cooper 1975). However, survival and regeneration rates from thermotherapy are very low, and plantlets regenerated from thermotherapy easily die following repeated subculture. Therefore, virus elimination using cryotherapy can be an ideal alternative for plants sensitive to elevated temperatures (Martin and Postman 1999). Cryopreservation is thought to facilitate viral elimination in larger meristems, as many viruses are unable to infect the meristem, and virus-free plants can be grown from excised apices of a critically small size (Martin and Postman 1999). Since Brison et al. (1997), many studies have reported that various clonal germplasms were free from serious virus infection following cryotherapy, and 50% of their cryopreserved cultures were rendered Plum Pox Potyvirus-free. Helliot et al. (2002) reported differences in the eradication frequency of Musa shoot meristems using cryotherapy for cucumber mosaic virus and banana streak virus. Similar findings (Kim et al. 2007) were reported for Allium using real-time quantitative PCR to cryogenically eliminate onion yellow dwarf virus, leek yellow stripe virus, and garlic virus A from unripe inflorescences.Fig.2
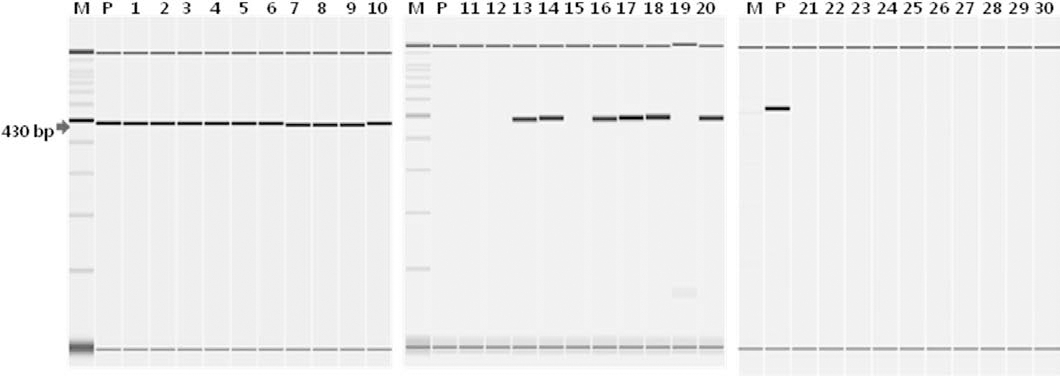
Fig. 2.
Detecting Potato leafroll virus (PLRV) by reverse transcription-polymerase chain reaction in virus-infected potato stock plants (Jasim) and plantlets regenerated following shoot tip culture and cryotherapy (encapsulation-vitrification). Nucleotides were the expected amplification products from PLRV. (M) 100-bp DNA ladder marker; (lanes 1~10) detecting PLRV before shoot tip culture; (lanes 11~20) detecting PLRV after shoot tip culture; (lanes 21~30) after cryogenic treatment (encapsulation-vitrification).Effects of shoot tip size used for cryotherapy on shoot recovery and virus eradication
Shoot tips of four different sizes (0.5, 1.0, 1.5, and 2.0 mm) were excised, and the rates of survival and regeneration were estimated using the encapsulation-vitrification method. As shown in Table 3, the highest eradication rate (98.3~99.1%) was found when the cryopreserved shoot tips were 1.0-1.5 mm in length and the frequency of PVY and PLRV eradication from regenerated plantlets was similar (96.4~99.3% and 97.3~ 99.7%, respectively), regardless of the shoot tip size. This result is very similar to that reported by Ding et al. (2008). Although shoot tip size can be a critical factor in conventional virus elimination, this may not be the case with cryotherapy. Wang et al. (2003) reported that 97% of grapevine virus A can be eradicated using vitrification and encapsulation-dehydration, independent of size and cryopreservation protocol. PVY and PLRV virus-free frequencies obtained by encapsulation-dehydration, encapsulation-vitrification, and droplets were the same (Wang et al. 2006). We evaluated the rates of regeneration and eradication between cryotherapy and meristem culture alone. As shown in Table 4, meristem tip regrowth rates increased with size. The 0.1 mm meristem tips did not survive to regenerate shoots. The 0.5 mm meristem tips regrew to 99.1% of the recovered shoots, but all plantlets grown in vitro remained PVY- and PLRV-infected as determined by RT-PCR. The best results for PVY (20.3%) and PLRV (23.2%) eradication and a 20.3% regeneration rate were found with the 0.3-mm meristem tips. This result is very similar to that reported by Ding et al. (2008). Meristem size is a very critical factor for determining virus-free frequency in meristem culture. Virus-free frequency is negatively related with meristem size in culture. Therefore, it is advisable to excise 0.3-mm-long meristem tips and culture them to eradicate PVY and PLRV, but the rates of eradication were much lower than those using the cryotherapy methods. We conclude that cryopreservation of shoot tips using droplet-vitrification or encapsulation-vitrification is an ideal option for producing PVY and PLRV-free potato stocks. Wang et al. (2006) reported similar results and produced much higher frequencies of virus-free plantlets than those derived from meristem culture and required a shorter production time for virus-free plants than when using other virus elimination methods. Because virus-free donor material is required for successful cryopreservation and conservation of accessions, Wang et al. (2006) proposed the use of cryopreservation for long-term storage of potato accessions and the production of virus-free plants.
Table 3.
Effect of shoot tip size on shoot recovery and eradication of Potato virus Y (PVY) and Potato leafroll virus (PLRV) from Solanum tuberosum cv. Jasim following cryotherapy.
| Size of explants (mm) | Survival rate (%) | Recovery rate (%) | Virus eradication rate (%) | |
|---|---|---|---|---|
| PVY | PLRV | |||
| 0.5 | 97 | 71.2±2.71b | 99.3±3.09a | 98.3±2.13a |
| 1.0 | 100 | 98.3±2.32a | 96.4±3.11a | 99.7±1.31a |
| 1.5 | 100 | 99.1±5.30a | 97.9±1.54a | 97.3±2.71a |
| 2.0 | 98 | 74.3±2.42b | 98.9±1.72a | 98.3±3.40a |
Table 4.
Effect of meristem tip size on shoot recovery and eradication of Potato virus Y (PVY) and Potato leafroll virus (PLRV) from Solanum tuberosum cv. Jasim following meristem culture alone.
| Size of explants (mm) | Survival rate (%) | Recovery rate (%) | Virus eradication rate (%) | |
|---|---|---|---|---|
| PVY | PLRV | |||
| 0.1 | 0 | - | - | - |
| 0.3 | 100 | 68.3±2.31b | 60.3±2.15a | 63.2±2.31a |
| 0.5 | 100 | 99.1±5.35a | 62.5±3.12a | 65.3±4.27a |
Preserving plant genetic resources is one of the cornerstones of breeding new cultivars for future needs, but germplasm collections should not inadvertently constitute a source of pathogens disseminated to new areas. Cryopreservation of potato germplasms is an alternative chemo- or thermotherapy method with the advantage that cryotreated shoot tips can be simultaneously stored for long-term conservation.
