

초롱꽃과(Campanulaceae)에 속하는 도라지(Platycodon grandiflorum) 는 한국, 일본, 중국 등에 분포하며 다년생의 직근 성 비대형 식물이다. 우리나라에서는 예로부터 도라지의 괴 근(塊根)을 채소 또는 약용으로 이용해 왔으며, 뿌리에는 saponin, inulin, phytosterin, platycodinin 등이 함유되어 있 어 용혈(溶血), 진해(鎭咳), 거담(祛痰) 및 해열(解熱) 등에 효과가 있는 것으로 알려져 있다.
경제 및 문화의 발달과 함께 예방의학과 건강에 대한 관 심이 커지면서 다양한 생리활성 성분을 함유한 유용식물로 부터 천연물 기능성 소재 발굴의 필요성 및 중요성이 부각 되고 있다. 특히 환경오염 및 공해에 의한 기관지의 질병이 증가 추세로, 도라지의 여러 가지 약리효과를 검증한 결과, 화학약품의 부작용에 대한 대안의 하나로 인정되어 예방적 측면, 치료적 목적으로 많이 이용되고 있다.
Proteomics는 기능성 단백질 해석, 단백질의 구조변화 게 놈의 기능, 생물기능을 해명하는 연구분야로, 최근의 proteome 이라는 용어는 특정 기관 혹은 조직에서 특정 스트레스에 발현되는 모든 단백질의 발현에도 적용되고 있다(Woo et al., 2003; Kamal et al., 2012).). 1990년대까지 특정 단백질 분석에 관련된 연구는 주로 MALDI-TOF MS기술을 이용 하여 단백질을 동정하는 방향으로 이루어져 왔으나, 2000 년대에 들어서는 복합단백질의 질량분석이 가능한 다차원 단백질 동정기술(MudPIT)과 구축되어 있는 database 및 생 물정보 관련 program 을 이용하는 bioinformatics를 접목하 는 방향으로 많은 연구가 수행되고 있다(Kim, 2010; Lee, 2009). 최근에는 복합 단백질의 질량분석이 가능한 다차원 적 단백질 동정기술(MudPIT=multi dimension proteinidentify technology)을 이용한 연구가 활발하다(Kamal et al., 2012; Link et al., 1999; Washburn et al., 2001). MudPIT는 SDSPAGE에서 발현된 복합단백질을 1차적으로 양이온으로 치 환시킨 후, 2차적으로 역상크로마토그래피에 의해 분리된 peptides를 ion-trap 사중극에서 전압에 의해 분리된 peptides 의 질량값을 구하고 database search를 통해 대량의 단백질을 동정하여 목적 단백질의 기능을 해석한다(Kim et al., 2010; Wolters et al., 2001). 대량의 동정된 단백질 bioinformation을 바 탕으로 단백질 사이에서의 네트워크 분석을 통해 식물체에 미치는 스트레스의 메카니즘을 분자 수준에서 해석하기 위 한 연구가 활발히 진행 중이다(Komatsu et al., 2013; Meunier et al., 2007). 따라서 본 연구는 2배체와 4배체 도 라지 단백질 발현지도 제작과 도라지의 생리활성에 관여하 는 메카니즘을 분자적 수준에서 해석하기 위한 기초자료를 얻고자 실시하였다.
재료 및 방법
공시재료
재배품종은 우송정보대학의 시험포장 비닐하우스에서 재 배되고 있는 2년생 재래품종 2배체와 4배체 도라지의 뿌리 를 분양받아 공시재료로 사용하였다(Fig. 1). 도라지의 뿌리 는 근부의 표피에 묻은 흙을 완전히 제거하기 위해 수회 세 척한 후, 박피하여 3차 증류수로 다시 세척하였다. 물기를 완전히 제거한 후, -80°C에서 보관하였다.
단백질 추출
단백질 추출에서 효과적인 세포벽 파쇄와 불순물 제거를 위해 액체질소와 TCA 침전법을 이용하여 실시하였다. 단 백질 추출을 위해 2배체와 4배체 도라지 뿌리 1g을 액체질 소로 마쇄한 뒤, 20 mM Tris-CI (pH 8.0), 0.2% SDS, 200 mM DTT의 조건하에서 10분간 중탕한 다음 상등액을 회수 하여 추출된 단백질에서 페놀계 화합물의 제거를 위하여 10% TCA로 -20°C에서 1시간 동안 단백질을 침전시켰다. 침전된 단백질들은 acetone에 용해시켜 3회 세척하였으며, 원심분리하여 상등액을 버린 후 speed vacuum centrifugation 으로 건조시켰다. 건조된 단백질에 lysis buffer [6M Urea, 2M Thiourea, 10mM Tris HCl (pH 8.0), 4% CHAPS, 50mM Dithiothreitol (DTT), 0.4% Protease Inhibitor Cocktail, 0.75% Ampholine (pH 3-10)] 300㎕을 추가하여 1시간동안 vortexing 한 후, 20분간 원심분리(20,000 rpm)하여 상층액을 새 e-tube 에 옮겨 RC/DC 분석에 의해 단백질 농도를 결정하였다.
2-Dimensional Electrophoresis
일차원 전기영동(one-dimensional electrophoresis: 1-DE) 은 등전점 전기영동으로, 15 cm 유리관에 straw gel (Urea, 30% acrylamide, 10% NP-40, Ampholine pH 3-10, 10% APS, TEMED)을 조성하였으며, 4시간 이상 굳힌 후, 70~150 μl의 시료를 주입하여 전기영동하였다. 전기영동 조건은 150V (1h), 300V (1h), 500V (16h)에서 비평형 전기영동하 였다. 일차원 전기영동 후 평형화가 끝난 겔을 이차원전기 영동을 하였다. 2차 전기영동 분석을 위해 12% separating gel과 5% stacking gel에 loading 시킨 후 Coomasie Brilliant Blue (CBB, R-250)로 염색하였다. 각각의 gel은 image 분 석을 위하여 scanner (HP Scanjet G4010, USA)로 스캔하였 으며 2D-gel image는 Progenesis samespot (Nonlinear Dynamics, Newcastle-upon-Tyne, UK)을 사용하여 분석하였다.
In-gel digestion
이차원전기영동 후 겔에서 원하는 spot을 적당한 크기(1 mm X 1 mm)로 잘라서 e-tube에 넣어준 다음 미리 준비해 둔 200 μl 증류수가 담긴 PCR tube에 넣어주었다. Vortexing 후, 증류수는 제거하고 gel band 세척을 위하여 30% methanol 1 ml를 첨가하여 재차 vortexing을 수행하였으며, 30% methanol 과정은 gel을 염색한 CBB가 탈색되어 gel이 하얗게 될 때 까지 반복하였다. 탈색 후, methanol은 제거하고 50% ACN/ 10mM ABC 200 μl을 첨가하여 gel을 추가적으로 세척해 준 다음 제거 후, 다시 100% ACN을 첨가하여 상온(25°C) 에서 10분간 반응시켜 gel을 탈수화 시켰으며, 반응이 끝나 면 ACN을 제거하고 speed vac.을 사용하여 완전히 건조시 켰다. 완전히 건조된 sample은 reduction reagent (10mM DTT in 100mM ABC) 50 μl를 첨가하여 56°C에서 1시간 반응시킨 다음, 2분간 원심분리(12,000 rpm)한 후 상등액을 제 거해 주었다. 이후 alkylation reagent (55mM IAA in 100mM ABC) 50 μl을 넣고 상온(25°C), 암조건 하에서 40분간 반 응시킨 후 원심분리하여 상등액을 제거하였으며, 증류수로 세척하였다. 세척과정 후, 증류수를 제거하고 100% ACN 10 μl를 첨가하여 다시 gel을 10분간 탈수화 하였으며, 이 후 ACN을 버리고 10분간 speed vac.으로 완전히 건조시켰 다. 완전히 건조된 sample은 digestion과정을 수행하기 위해 건조된 sample에 trypsin enzyme 50 μl 37°C에서 16시간 반응시켜 digestion 시킨 후 건조 시켰다.
질량분석 및 bioinformatics를 이용한 단백질 동정
In-gel digestion과정을 통해 분해된 peptide를 동정하기 위한 모든 MS/MS분석은 surveyor HPLC system과 nano-ESI 소스가 장착된 7-tesla Finningan LTQ-FT-ICR mass spectrometer LTQ-FT-ICR 질량 분석기(Therm Electron, Bremen, Germany) 로 구성된 Nano-LC/MS 시스템으로 수행되었다. MS분석은 먼저, sample을 탈염(脫鹽)화 시키고 20 μl/min의 flow rate로 농축하기 위해 각각의 sample 10 μl을 auto sampler (Surveyor) 를 사용하여 C18 trap column (I.D., 300 μm, length 5 mm, particle size 5 μm, LC Packings)에 주입하였으며, 이후 자 동적으로 MS와 MS/MS acquisition을 반복하며 data-dependent mode로 작동하였고, 다음과 같은 조건하에 수행되었다 (Conditioon: 2.2kV spray voltage, 220°C ion transfer tube temperature, 1.3millitorrs collision gas pressure). MS/MS acquisition에서 ion 검출은 500 counts를 한계점으로 설정 하였으며, 활성화는 q=0.25, 활성화시간은 30ms로 설정하 였다. LTQ-FT-ICR MS로 동정된 peptide의 MS spectra data값은 먼저 MASCOT ver.2 (Matrix Science, UK)의 database에서 검색하였으며, Olsen & Mann (2006)에 의거 하여 peptide 오차범위는 50 ppm, fragment ion 오차범위는 0.5 Da로 설정하였다. 수집된 protein description과 peptide sequence의 정보는 (http://pir.georgetown.edu)을 통해 protein sequence를 찾는 검색기반으로 사용하였다. 최종 확인된 도 라지 단백질은 분자량, pI 2가지 항목을 통해 각각의 특성 에 따라 분류하여 도라지 단백질의 일반적인 화학적 특성을 파악하였고, 이후 도라지 단백질의 protein sequence을 이용 하여 UniProt을 통해 단백질 기능을 확인하였으며, 확인된 기능별로 나누어 단백질을 분류하였다.
결과 및 고찰
2배체와 4배체 도라지뿌리의 단백질 발현 양상
이차원 전기영동을 실시하여 2배체 및 4배체 도라지 뿌 리로부터 추출한 단백질의 발현 양상은 Fig. 2, 3과 같다.
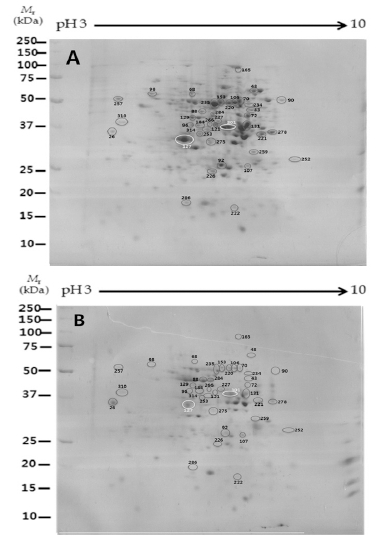
Fig. 2.
2-D patterns of proteins expression from (A) diploid and (B) tetraploid root. Proteins were separated on a 2-D gel using 11 cm tube pH 3-10 L in the first dimension and 2D-PAGE in the second dimension.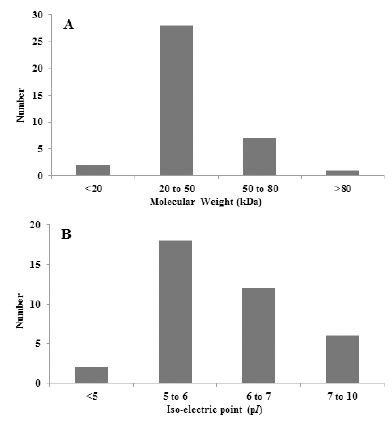
Fig. 3.
Frequency distribution for molecular weight and isoelectric point of identified proteins from diploid and tetraploid root. (A) Molecular weight (kDa), (B) Iso-electric point (pI).발현된 단백질 spot을 Progenesis SameSpot software로 이 미지 분석하여 찾은 535개의 단백질 spot 중(≤ 2-fold)인 단백질 67개를 선택하였다. 이들 중 서로 다른 양상을 보이 는 단백질 39개를 최종 선택하여 mass spectrometry 질량분석 기로 분석·동정을 한 spot은 원으로 표시하였으며 numbering하 여 나타내었다. 2배체 및 4배체 도라지의 단백질의 발현 양 상은 배수성에 따른 특이성은 관찰 할 수 없었으며, 모두 분 자량 15~100 kDa 크기, pH 4.0~8.0의 범위에 분포하는 것 을 볼 수 있었다(Fig. 2). 특히, 분자량은 20~50 kDa 범위에 서 약 74% 정도로 가장 많은 단백질이 분포하였으며, 중분 자 범위의 50~80 kDa도 약 18% 정도 분포하는 것으로 나 타났다. 등전점(pI) 또한 배수성에 관계없이 pI 5~6 사이에 서 약 47%로 가장 많은 단백질 분포를 보였으며, pI 6~7사 이의 단백질 경우도 약 29% 정도 분포하여, pI 7 이하의 단 백질이 80% 이상으로 나타났다(Fig. 3). 이 같은 결과로부 터 도라지 뿌리의 경우, 배수성에 관계없이 산성 단백질이 많은 것을 알 수 있었다. 애기장대는 대부분 작은 분자량을 가진 단백질로, 10~50 kDa의 분자량을 가지는 단백질이 가 장 많았으며, 염기성 단백질이 많이 포함되어 있다(Millar et al., 2005)고 하였으나, 도라지 뿌리의 경우 분자량에 따 른 분포는 애기장대와 유사한 결과를 보였으나, 산성 단백 질이 많이 포함되어 차이가 있음을 보였다.
2배체와 4배체 도라지 뿌리의 프로테옴 동정
LTQ-FT-ICR MS를 이용하여 2배체와 4배체 도라지 뿌 리에서 총 39개의 단백질이 동정되었으며, 이들의 농도 및 기능을 분류한 결과는 Fig. 4와 Table 1과 같다. 동정되었던 각 단백질의 농도는 2배체의 경우, 대장균 L10의 리보좀단 백질과 기능적으로 같으며, 리보좀단백질 L10P과(科)에 속 해 단백질 합성의 신장단계에 중요한 역할과 대부분의 리보좀 단백질이 염기성인 것과는 달리 산성인 60S acidic ribosomal protein P0 (SN 127; 34.4 kDa, pI 5.38) (Kenmochi et al., 1998), 산화환원효소의 활성과 관련있는 단백질(SN 131; 39.1 kDa, pI 6.86), 해당작용·알코올발효에 있어 고에너지 인산결합을 가지는 화합물의 생성에 중요한 반응을 촉매하 는 효소로서 생물계에 널리 분포하는 Enolase (SN 235; 47.9 kDa, pI 5.92) 및 구연산회로에서 malate를 OAA (oxaloacetic acid)로 또는 그 반대의 화학반응을 촉매하는 산화한원 효 소로, 당의 생성에도 관여하는 malate dehydrogenase (SN 301; 35.7 kDa, pI 6.33) (Goward et al., 1994)에서 높은 것 으로 나타났다. 반면, 4배체는 당의 분해와 생성에 중요한 효소이며, 1,3-bisphosphoglycerate에 glyceraldehyde 3-phosphate 와 무기인산염의 가역적 전환의 촉매작용 역할을 하는 glyceraldehyde-3-phosphate dehydrogenase (SN 26; 36.9 kDa, pI 7.7)와 해당과정에 있어 중요한 효소인 phosphoglycerate kinase (SN 88; 42.3 kDa, pI 5.69)의 농도가 높았 다. 그러나 SN 26과 SN 88을 제외한 모든 단백질에서 4배 체에 비해 2배체에서 높은 농도를 가지는 것으로 나타났다. 특히 2배체의 SN 127, SN 131, SN 235 및 SN 301의 단백 질은 4배체보다 2배 이상의 높은 농도를 보였다(Fig. 4, Table 1). MASCOT search engine을 통해 얻어진 정보를 바탕으로 spot number (SN), protein accession (PA), 단백 질 이름(Protein description), 단백질 기능(proteion functions), molecular weight (MW), iso-eletric point(pI), protein matches (PM), protein coverage (PC), peptide sequence (PS)등으로 분류한 결과는 Table 1, Fig. 5과 같다.

Fig. 4.
Relative protein intensity of differentially expressed proteins from diploid and tetraploid root.Table 1.
List of identified proteins from diploid and tetraploid roots in Platycodon grandiflorum using LTQ-FT-ICR MS/MS.
| SN1) | PA2) | Protein Description | MW3) | pI4) | PM5) | PC6) | PS7) |
|---|---|---|---|---|---|---|---|
| Oxidoreductase activity | |||||||
| 26 | P04796 | Glyceraldehyde-3-phosphate dehydrogenase | 36901 | 7.7 | 6 | 29.6 | IGINGFGR |
| 48 | Q43839 | Glucose-6-phosphate 1-dehydrogenase | 65646 | 6.88 | 8 | 15.8 | IDHYLGK |
| 70 | P81406 | NADP-dependent glyceraldehyde-3-phosphate dehydrogenase | 52970 | 6.76 | 4 | 6.2 | IAWEEPFGPVLPVIR |
| 72 | Q9S7E4 | Formate dehydrogenase | 42383 | 7.12 | 1 | 4.2 | DWLESQGHQYIVTDDK |
| 90 | Q9SNX8 | NADP-dependent glyceraldehyde-3-phosphate dehydrogenase | 53139 | 7.49 | 16 | 22.8 | DSVTEVVR |
| 131 | Q7XWU3 | Probable cinnamyl alcohol dehydrogenase6 | 39061 | 6.86 | 3 | 11.1 | YRFVINVGGDSK |
| 221 | Q43247 | Glyceraldehyde-3-phosphatedehydrogenase | 36426 | 7.01 | 7 | 11.3 | TLLFGEK |
| 259 | P52578 | Isoflavone reductase homolog | 33831 | 6.16 | 6 | 17.2 | VHAVEPAK |
| 278 | P26521 | Glyceraldehyde-3-phosphate dehydrogenase | 36547 | 7.66 | 31 | 26 | VLPALNGK |
| Ion binding | |||||||
| 43 | P50217 | Isocitrate dehydrogenase [NADP] | 46762 | 6.54 | 11 | 20.7 | WPLYLSTK |
| 235 | Q9LEI9 | Enolase2 | 47884 | 5.92 | 39 | 24 | EGLELLK |
| 257 | Q38858 | Calreticulin-2 | 48127 | 4.37 | 6 | 17.7 | LDCGGGYMK |
| 284 | P25696 | Bifunctional enolase 2/transcriptional activator | 47689 | 5.54 | 5 | 16.2 | TYDLNFK |
| Nucleotide binding | |||||||
| 68 | P37399 | ATP synthase subunit beta, mitochondrial | 59099 | 5.62 | 5 | 11.5 | AEKMAKENPQ |
| 88 | Q42962 | Phosphoglycerate kinase | 42338 | 5.69 | 5 | 14.2 | IGVIESLLEK |
| 98 | P21239 | RuBisCO large subunit-binding protein subuni talpha | 57657 | 4.84 | 5 | 8.1 | LAGGVAVIK |
| 106 | P12782 | Phosphoglycerate kinase | 49809 | 6.58 | 4 | 10.6 | SLAFAAGADPR |
| 129 | Q05214 | Actin | 41712 | 5.46 | 10 | 26 | RGILTLK |
| 165 | P46523 | ATP-dependent Clp protease | 97259 | 5.9 | 2 | 4.7 | VAVGNNVLDTLGRSR |
| Peptidase activity | |||||||
| 92 | Q8L4A7 | Proteasome subunit alphatype-2-B | 25717 | 5.53 | 3 | 23.4 | LYKEPIPVTQLVR |
| 107 | Q7DLS1 | Proteasome subunit beta type-7-B | 29598 | 6.71 | 5 | 10.6 | DGVILGADTR |
| Transferase activity | |||||||
| 96 | Q42653 | Quercetin 3-O-methyltransferase2 | 37843 | 5.68 | 5 | 14 | GMSDHSTITMK |
| 184 | O81646 | Caffeic acid3-O-methyltransferase | 39610 | 5.62 | 8 | 28.7 | GINFDLPHVIR |
| 220 | P19595 | UTP--glucose-1-phosphateuridylyltransferase | 51841 | 5.7 | 7 | 17.4 | LDALLAKGK |
| 266 | O81646 | Caffeic acid3-O-methyltransferase | 39610 | 5.62 | 4 | 12.3 | GINFDLPHVIR |
| 275 | O23733 | Cysteine synthase | 33881 | 5.67 | 4 | 12.7 | LEMMEPCSSVK |
| 275 | O23733 | Cysteine synthase | 33881 | 5.67 | 4 | 12.7 | LEMMEPCSSVK |
| Catalytic activity | |||||||
| 121 | O82560 | Glutamine synthetase cytosolic isozyme 2 | 39184 | 6.12 | 5 | 15.4 | EHIAAYGEGNER |
| 227 | P07694 | Glutamine synthetase root isozyme A | 39264 | 6.12 | 3 | 13.7 | DIVDAHYK |
| 301 | P57106 | Malatedehydrogenase | 35652 | 6.33 | 2 | 12 | MELVDAAFPLLK |
| Structural molecule activity | |||||||
| 127 | P41095 | 60S acidic ribosomal protein P0 | 34356 | 5.38 | 3 | 14.7 | GTVEIITPVELIK |
| 253 | O22518 | 40S ribosomal protein SA | 33885 | 5.1 | 4 | 14.5 | LLILTDPR |
| Metal cluster binding | |||||||
| 153 | O22553 | Cholinemonooxygenase | 50187 | 6.02 | 3 | 11.4 | VAEWGPFILISLDR |
| Ligase activity | |||||||
| 222 | Q9CAB6 | Ubiquitin-conjugatingenzyme | 17989 | 5.03 | 2 | 22.6 | LLEELER |
| Conjugating protein ligase activity | |||||||
| 234 | Q9LT79 | U-boxdomain-containingprotein 25 | 46027 | 7 | 3 | 4 | AAALR |
| Transporter activity | |||||||
| 252 | P42056 | Mitochondrial outer membrane protein porin of 36 kDa | 29376 | 7.78 | 8 | 14.5 | IGLAVALKP |
| Hydrolase activity | |||||||
| 310 | Q9ZPF5 | Probablearginase | 37957 | 5.9 | 2 | 9 | FGVEQYEMR |
| 314 | Q9SB68 | Manganese-dependent ADP-ribose/CDP-alcohol diphosphatase | 34715 | 5.82 | 1 | 5.1 | VIVCGHVPMSPGVASK |
| Response to stimulus | |||||||
| 286 | P04793 | 17.5kDaclassIheatshockprotein | 17535 | 5.33 | 2 | 13.1 | FRLPENAK |
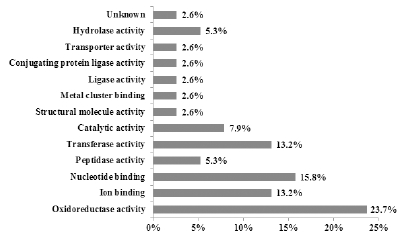
Fig. 5.
Functional classification of identified proteins from diploid and tetraploid roots of Platycodon grandiflorum.2배체와 4배체 도라지 뿌리의 단백질에는 생체에 있어 많은 유기화합물 및 상당수의 무기화합물의 산화환원 반응 을 촉매하여, 생활과 생체구성성분의 합성에 필요한 에너지 를 얻는 역할을 하는 효소로 알려진 산화환원효소의 활성 (oxidoreductase activity)기능을 갖는 단백질(SN 26, 48, 70, 72, 90, 131, 221, 259, 278)이 23.7%의 비율로 가장 많았 다. 다음으로 nucleotide binding기능의 단백질(SN 68, 88, 98, 106, 129, 165)이 15.8%의 비율로 높았으며, 특히 ATP synthase subunit beta (SN 68)는 엽록체 ATP synthase의 다섯 개의 소단위체 구성 요소중 하나다(Fritsche & Junge, 1996). 엽록체 ATP synthse는 틸라코이드막에 존재하는 두 개의 기능적 구성물인 CF0과 CF1을 가진 복합체로 CF0는 양성자의 이동통로로 소단위체의 구성, 구조, 기능이 미토 콘드리아의 F0와 유사한 막주변 단백질 복합체(peripheral membrance protein complex)이며 CF1은 여러개의 막에 고 정된 단백질로 ATP 합성장소이다(Grossman et al., 1995). Glyceraldehyde-3-phosphate dehydrogenase (SN 26)는 광 합성 중 탄소동화의 중요한 요소이며, 광합성 암반응 Calvincycle에 작용하는 효소이다. Peroxidase 단백질 효소는 화학 적 작용으로 독서의 산화제 및 환원제를 제거하며, 또한 환 경적인 스트레스에서의 반응은 상처, 병원균의 침투에 반응 하여 리그린과 코르크층 합성에 관여한다(Schweizer et al., 1989).
또한 transferase activity 기능의 단백질(SN 96, 184, 220, 266, 275), Ion-binding 기능을 갖는 단백질(SN 43, 235, 257, 284) 및 peptidase activity 기능의 단백질(SN 92, 107) 이 동정되었다. 기타, 물질이 에너지를 얻어 화학적으로 활 성되어 반응속도가 빨라지고 기능이 높아지는 촉매활성 (catalytic activity)기능의 단백질(SN 121, 227, 301)도 약 7.9% 정도 나타났다. 특히, malate dehydrogenase (SN 301) 는 구연산회로에서 malate를 OAA (oxaloacetic acid)로 또 는 그 반대의 화학반응을 촉매하는 산화환원 효소로, 당의 생성에도 관여하는 효소로 알려져 있다(Goward et al., 1994). 그밖에 structural molecule activity기능의 단백질(SN 127, 253), hydrolase activity (SN 310, 314), metal cluster binding (SN 153), ligase activity (SN 222), transporter activity (SN 252) 기능의 단백질 등이 동정되었다.
적 요
본 연구는 2배체와 4배체 도라지 단백질 발현지도 제작 과 도라지의 생리활성에 관여하는 메카니즘을 분자적 수준 에서 해석하기 위한 기초자료를 얻고자 실시하였다. 2배체 및 4배체 도라지의 단백질의 발현 양상은 배수성에 따른 특 이성은 관찰 할 수 없었으며, 모두 분자량 15~100 kDa 크 기, pH 4.0~8.0의 범위에 분포하는 것으로 나타났다. 동정 된 39개의 단백질 중 2배체에 비해 4배체에서 2개의 단백 질이 up-regulated 되었고, 37개의 단백질이 down-regulated 되었다. 단백질을 기능별로 분류한 결과, 산화환원효소의 활성(oxidoreductase activity)기능을 갖는 단백질이 23.7% 의 비율로 가장 많았고 다음은 nucleotide binding 기능의 단백질이 15.8%의 비율로 높았다.
