

서 론
재료 및 방법
시험재료 및 고구마의 농업적 특성조사
엽록소 형광 및 엽록소 함량 측정
당도 분석
통계분석
결과 및 고찰
차광량별 처리에 따른 1일 광량의 비교
생육초기 차광정도에 따른 잎의 엽록소 형광 반응
생육초기 차광정도에 따른 잎의 엽록소 함량
생육시기별 차광처리 직 후 생육 및 괴근 형성
생육시기별 차광 정도에 따른 수확 후 괴근 수량 및 괴근 당도
적 요
서 론
고구마(Ipomoea batatas L.)는 아열대 작물로서 일반적으로 일년생 작물로 재배된다. 재배조건과 품종에 따라서 재배기간은 12주와 35주 사이로 다양하다(Hahn & Hozyo, 1984). 광은 식물의 광합성과 형태발생에 관여하는 필수적인 요소 중 하나로서 작물의 수량에 결정적인 영향을 미친다. 또한 수량 감소는 바이오 매스 생산 감소 및 식물체 내에서 변경된 동화 분배로 인해 발생하고, 괴근 발생보다는 괴근 형성에 영향을 미쳤다고 보고되었다(Oswald et al., 1995). 고구마의 괴근 및 총 바이오 매스 수량이 차광처리에 의해 품종에 따라 높은 유전적 다양성을 보였으며, 차광에 내성인 품종은 모든 성장 단계에서 거의 영향을 받지 않은 반면 민감한 품종은 20%에서 55% 사이의 수확량 감소를 나타냈다. 일반적으로 괴근 수량 감소는 생육 후기에 일사량이 부족 할 때 가장 심했고 생육초기에 일사량이 부족했을 때는 영향을 덜 받는다고 보고되었다(Oswald et al., 1995). 토양이 심하게 건조하지 않은 조건에서 고구마 재배는 일조량이 높은 것이 좋고(약 11시간), 일조량이 부족하면 광합성이 감소할 뿐 아니라 지상부에서 생산된 물질이 지상부에 이용되고 덩이뿌리에 저장되는 양이 적기 때문에 지상부의 줄기와 잎 수량은 증가하나 고구마수량은 크게 감소한다고 하였다(Jeong et al., 2003; Han et al., 2012).
최근 잦은 태풍 및 긴 장마기간으로 인한 일조부족이 문제되어 작물의 수량 감소 피해가 발생하고 있다. 국내에서는 주로 벼의 차광처리에 의한 생육 및 수량특성 관련 연구가(Yang et al., 2007; Kim et al., 2014; Lee et al., 2014; Lee et al., 2016) 다수 보고되어 있으나, 현재 고구마 생육시기별 일조량 부족에 따른 피해양상 및 피해율에 대한 연구는 미미한 실정이다. 이에 본 연구는 생육시기에 따라 차광량을 달리하여 고구마의 생육양상 및 수량 감소 정도를 구명하고자 수행하였다.
재료 및 방법
시험재료 및 고구마의 농업적 특성조사
본 시험은 국립식량과학원 바이오에너지작물연구소 시험연구포장에서 수행되었다. 시험품종은 점질 고구마 ‘호감미’와 분질 고구마 ‘진율미’를 고구마 표준재배법에 준해 2019년 5월 13일에 정식하여 재배하였다. 정식간격은 이랑넓이 70 cm, 주간간격은 20 cm로 재배하였으며, 차광처리는 55%, 75%로 시판용 차광망을 사용하여 철골하우스 형태에 차광량별로 설치하여 처리하였고 무처리구를 대조구로 조사하였다. 철골하우스의 크기는 가로 6 m, 세로 12 m로 설치하였고, 시험구는 난괴법 3반복으로 배치하였다. 1일 광량은 Lighter meter (Lutron) LX-1128SD 데이터 로거를 설치하여 1시간 간격으로 1일중 09시부터 19시까지 모니터링 하였다. 처리 시기는 괴근형성기(정식~정식 후50일), 괴근비대기(정식 후 50~90일), 괴근비대성기(정식 후 90~120일)로 처리하였다. 생육 시기별 차광 처리 직 후 주당 10 g 이상 괴근 형성 수 및 무게, 지상부‧지하부 건물중, T/R율, 엽면적을 조사하였고 수확 후(120일 후) 괴근 수량을 각 시험구당 3반복 씩 총 9개 샘플로 조사하였다.
엽록소 형광 및 엽록소 함량 측정
휴대용 엽록소 형광반응 측정기 Chlorophyll Fluorometer (OPTI-SCIENCES) OS1P를 사용하여 고구마의 정단부에서 엽폭이 5 cm 이상인 첫 번째 잎을 선정하여 총 15반복으로 엽록소 형광을 측정하였다. 생육 시기별 차광량 처리에 따른 Fv/Fm (광계 II 암적응 최대양자수율)을 측정하기 위해 30분 동안 암적응 후 측정하였다.
고구마 잎의 엽록소 함량은 총 6반복으로 95% (v/v)에탄올에 80°C에서 20분 동안 추출하였다. UV-Vis Spectrophotometer (Biochrom Libra S22)를 이용하여 648 nm과 664 nm 파장에서의 흡광도를 측정한 후 엽록소 A와 B 함량을 산출하였다(Vernon, 1960).
당도 분석
당도는 처리구별 9반복으로 찐고구마 20 g에 80 ml의 물을 넣고 믹서기로 분쇄 후 상온에서 굴절당도계(PR-32α refractometer, ATAGO,. Tokyo, Japan)를 이용하여 측정하고 희석배수 5를 곱하였다. 수분함량은 잘게 자른 생고구마 100 g을 80°C에서 예비 건조 한 다음 105°C에서 6시간 열풍 건조 한 후 건물 무게를 측정하여 100 g에서 뺀 값을 %로 나타내었다.
통계분석
모든 실험은 SPSS version 20 software (IBM, Chicago, IL, USA)를 이용하여 p<0.05, p<0.01, p<0.001 수준에서 분산분석(two-way or three-way ANOVA)과 p<0.05 수준에서 Tukey의 다중범위 검정으로 시료간의 유의성을 검정하였고, 데이터는 6~15반복의 평균값으로 나타내었다.
결과 및 고찰
차광량별 처리에 따른 1일 광량의 비교
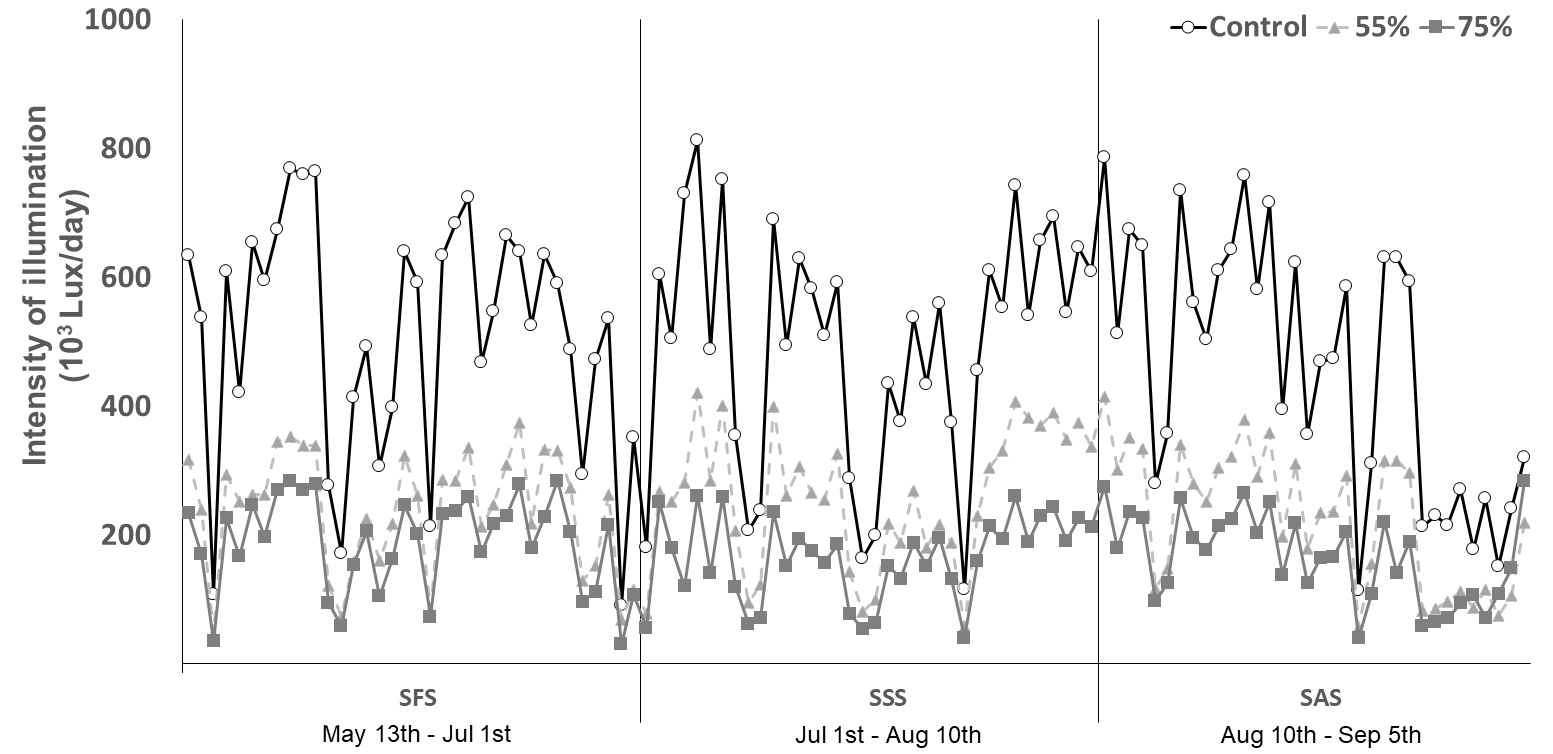
고구마 생육 시기별 차광처리에 따른 1일 광량을 9시부터 19시까지 모니터링 한 결과 괴근이 형성되는 생육초기(SFS;정식~50일, 5.13~7.1)에 차광처리 기간 동안은 대조구(자연광)의 광량(최대 812 klux, 최소 90 klux)에 비해 일평균 광량이 55% 차광처리는 52.9%, 75% 차광처리는 63.7%가 감소되는 수준을 보였다(Fig. 1). 그리고 괴근비대기(SSS:정식 후 50~90일, 7.1~8.10) 차광처리구의 광량은 대조구(자연광)에 비해 55% 차광처리는 47.9%, 75% 차광처리는 66.7%의 감소를 보였다. 마지막으로 괴근비대성기(SAS:정식 후 90~120일, 8.11~9.9)에는 55%와 75%차광처리구의 광량이 각각 52.4%, 63.3% 감소되었다(Fig. 1). 이를 통해 시판용 55%와 75% 차광막의 실제 차광률은 각각 평균적으로 약 51%와 65%로 약간 낮게 나타내는 것을 알 수 있었다.
생육초기 차광정도에 따른 잎의 엽록소 형광 반응
광합성 과정 중에 불합리한 환경 조건에 처하게 되면 광화학반응으로 흡수되는 에너지와 전자전달 반응에 이용되는 에너지 간에 균형이 깨지게 되는데 엽록소 형광분석은 이러한 현상에서 야기된 광합성의 상태 변화를 정량적으로 보여준다(Oh & Koh, 2004; Hopkins & Huner, 2008; Kang et al., 2016; Lee & Lee, 2017).
초기광화학반응에서 광계 II 암적응 최대양자수율(Fv/Fm)은 식물 잎이 광합성을 수행할 수 있는 최대값(잠재력)을 의미한다. 차광처리 50일 경(괴근형성기)에 차광정도에 따라 고구마 잎의 광합성 명반응 시 최대양자수율을 측정한 결과 호감미와 진율미 두 품종 모두 대조구와 55% 차광 시험구는 유의적인 차이를 보이지 않았고 75% 차광 시험구의 경우 대조구보다 약간 높은 수치를 보였다(Fig. 2A). 또한 괴근비대기인 차광처리 90일경에 광합성 최대 양자수율을 조사한 결과도 비슷한 결과를 보였다(Fig. 2B). 보통 광도가 낮은 환경에서 순화된 식물들은 광량을 많이 받기 위해 잎 내 질소를 엽록소 단백질에 공급하여 틸라코이드(thylakoid)내 광 집적 반응에 집중함으로써 전자전달계 단백질이나 루비스코(rubisco) 효소로 질소 공급이 상대적으로 줄어들어 전체적인 광합성 기능이 저해되기도 하지만 한편으로는 적응기작으로 광합성 양자수율은 높이고 호흡에 의한 에너지손실을 극소화시킴으로써 열악한 환경 조건에 적응하려는 반응을 나타내기도 한다(Ellsworth & Reich, 1992; Kim & Lee, 2001). Kang et al. (2016)의 연구에 의하면 차광처리에서 감국은 광계II 암적응 최대양자수율은 변화가 없었지만 광계I 전자전달 에너지 흐름이 대조구에 비하여 차광처리구(50%, 80%)에서 각각 14%, 43%씩 감소하여, 차광량이 높을수록 광이용효율이 저조해지는 것으로 나타났다. 꼬리풀, 꽃범의꼬리, 뱀무 및 약모밀의 경우도 이와 비슷한 경향을 보이는 것으로 보고되었다. 이러한 결과를 비추어 볼 때 고구마도 차광량 증가에 따라 광계II 암적응 최대양자수득률의 변화보다는 광계I 전자전달 에너지 흐름이 감소하여 광이용 효율이 감소하였을 것으로 추측된다.
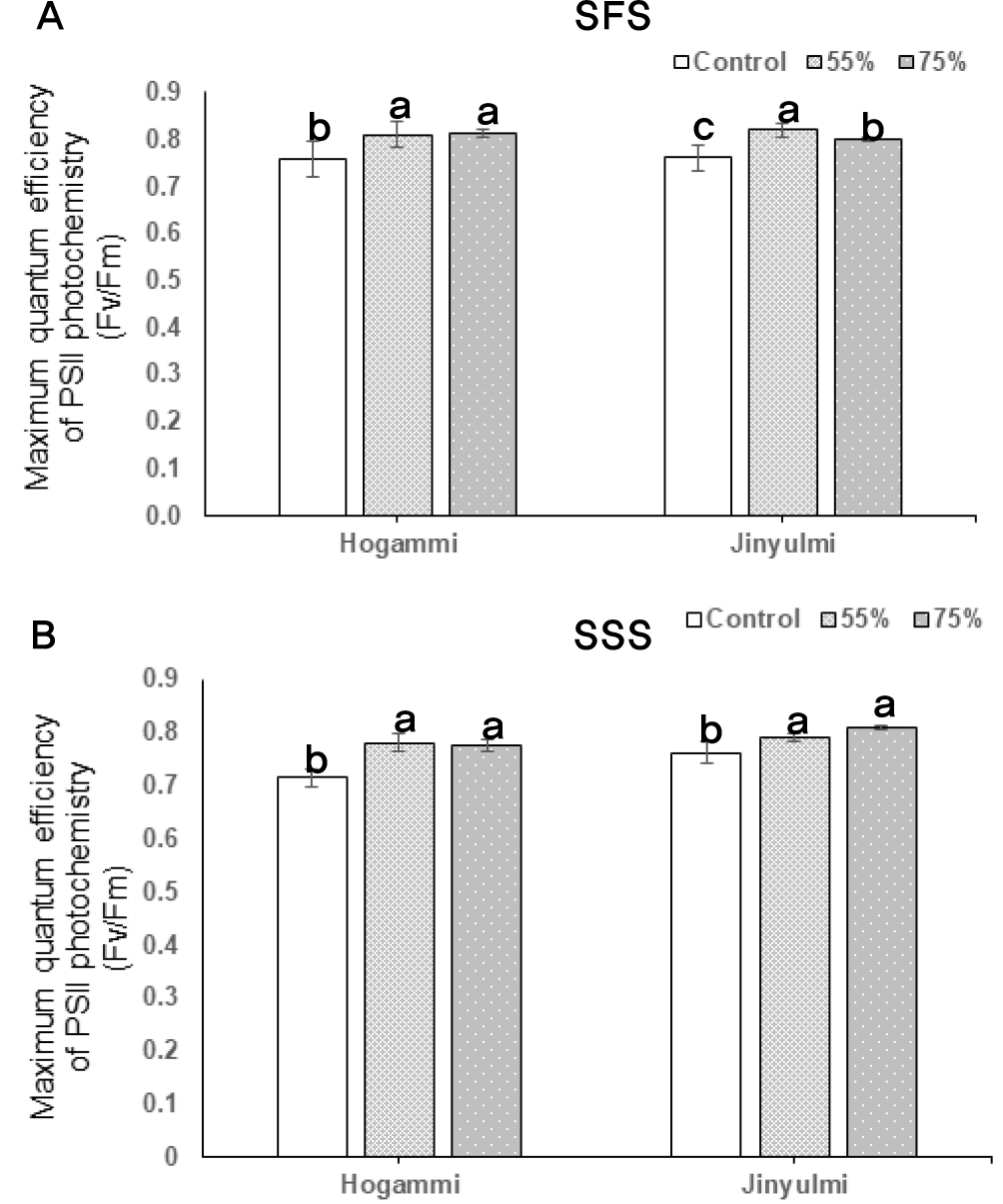
Fig. 2
Effects of shading on the maximum quantum efficiency of photosystem II photochemistry in sweetpotato. Sweetpotato plants were grown under different shading treatments (0%, 55%, and 75%). Chlorophyll fluorescence parameters of the plants were measured at the different storage root growth stages. SFS: storage root formation stage (planting~50th day), SSS: storage root swelling stage (50–90th day). Values are the averages of three replicates ± SD. Different lowercase letters in the figure indicate significant difference at P < 0.05, as determined using Tukey’s test.
생육초기 차광정도에 따른 잎의 엽록소 함량
차광 처리 50일 경과 후 처리별 광합성에 이용되는 색소의 함량을 조사하였다(Fig. 3). 호감미와 진율미 두 품종 모두 차광처리구의 엽록소 a와 b의 함량이 대조구에 비하여 유의적으로 증가하는 경향을 나타내었다. 호감미의 경우는 55%와 75% 차광처리 시 대조구에 비하여 엽록소a의 함량이 각각 9%, 10% 증가하였고, 진율미의 경우는 각각 3%, 7% 증가하였다. 엽록소 b의 함량은 특히 더 많은 비율로 증가 하였다. 호감미의 경우 대조구에 비하여 55%와 75% 차광처리구에서 각각 47%, 41% 증가하였고 진율미의 경우는 각각 39%, 34% 엽록소함량이 증가하였다. 이러한 결과는 차광막에 의하여 빛이 부족한 조건에서 광 수용능력을 유지하고자 식물이 엽록소 함량을 높은 수준으로 증가시키려고 하는 반응 때문인 것으로 생각되며, 수리취, 산마늘, 곰취 등 다른 식물들의 연구 결과에서도 비슷한 경향을 보여주었다(Lee et al., 2012c; Kim et al., 2015). 또한 식물이 뜨거운 열과 강한 광 조건에 장기적으로 노출되면 광합성 산물의 소비가 커지고 생육감소를 야기할 수 있다(Kim et al., 2017). 무차광 조건하에서 빛의 흡수를 줄이기 위한 광보호작용(Kyparissis et al., 2000; Lee et al., 2012a)으로 인해 엽록소 함량이 감소하기도 한다. 또 다른 원인으로는 차광 처리 시 대조구에 비해 잎의 노화를 지연시켜 엽록소 함량을 높게 유지하려고 하는 특성을 고려하기도 한다(Lee & Won, 2007). 이와 같이 차광 처리에 따른 엽록소 함량의 차이는 생육단계별 조사를 통한 추가 실험을 실시하여 정확한 원인을 구명할 필요가 있다. 대체적으로 광이 부족한 조건에서는 보다 많은 빛을 수용하기 위한 기작으로 반응중심 색소인 엽록소 a보다 광수확색소인 엽록소 b가 더 증가 되며, 엽록소 a/b가 감소한다고 알려져 있다(Lee et al., 2006; Lee et al., 2012c, 2012b; Kim et al., 2015). 고구마의 경우에도 차광처리시 대조구에 비하여 엽록소의 함량이 상대적으로 높게 나타났다. 하지만 식물 종에 따라 광 환경에 대한 반응이 다양하게 나타나는 것으로 판단하기도 하였다(Song & Kim, 2017).
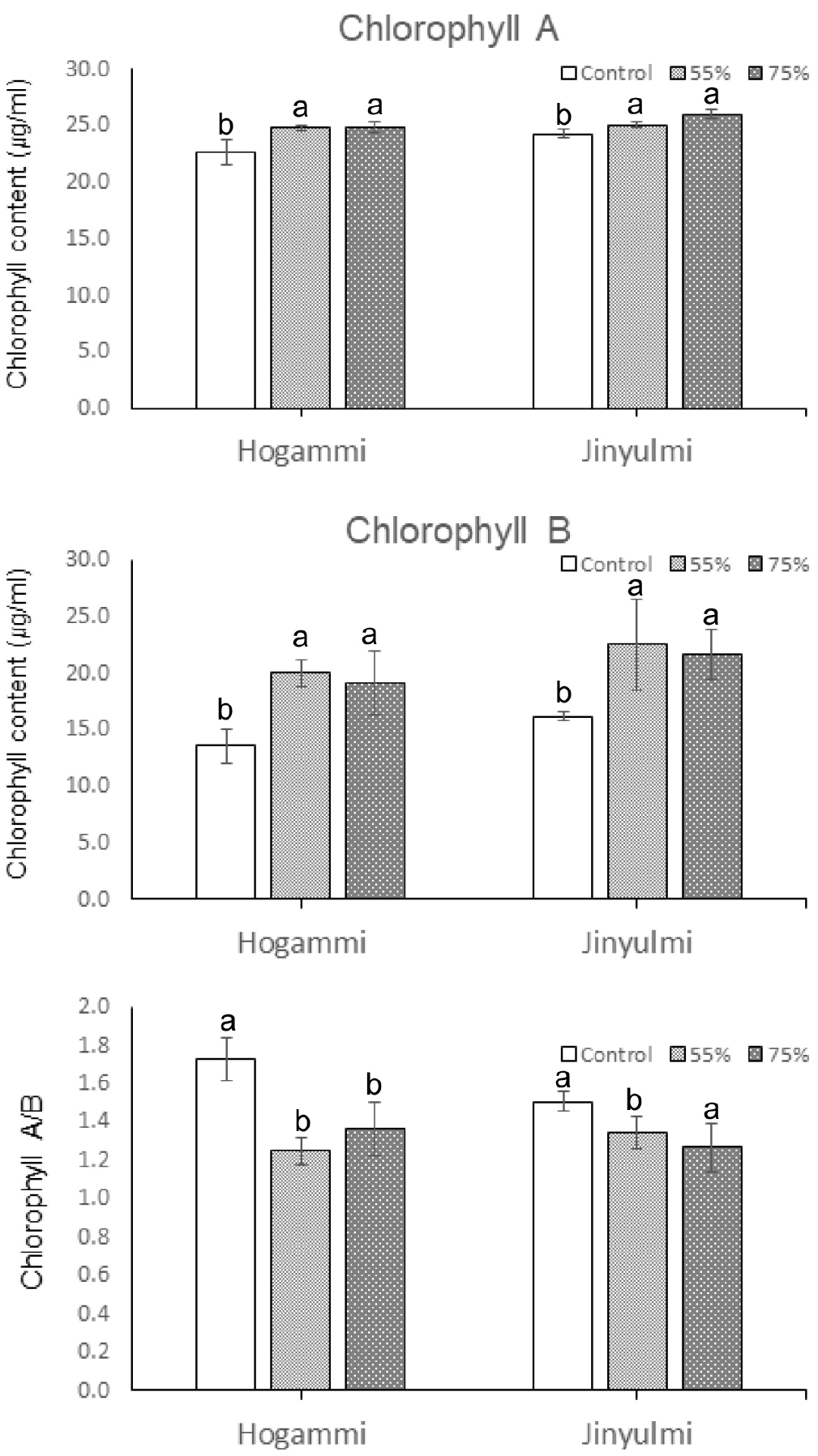
Fig. 3
Effects of shading on the pigment contents in sweetpotato. Sweetpotato plants were grown under different shading treatments (0, 55%, and 75%). The pigment content in plants were measured on the 50th day after commencing shading treatment. Values are the averages of three replicates ± SD. Different lowercase letters in the figure indicate significant differences at P < 0.05, as determined using Tukey’s test.
생육시기별 차광처리 직 후 생육 및 괴근 형성
괴근형성기(정식 후 50일간)동안 차광처리 직 후 주당 10 g 이상 괴근 형성 수 및 무게, 지상부‧지하부 건물중, T/R율, 엽면적을 조사하였다(Table 1, Fig. 4). ‘진율미’ 품종의 경우 10 g 이상 괴근 수는 대조구에서 주당 4.6개로 평균 18.3 g 수준 이었으나 55%와 75% 차광처리구의 경우 주당 괴근 개수는 각각 평균 0.2 g과 0.1개로 감소하였으며, 주당 괴근 무게는 각각 평균 3.2 g과 1.5 g 수준이었다. ‘호감미’ 품종도 역시 10 g 이상 괴근수는 대조구 3.2개, 평균 괴근중 10.6 g이었으나 55%와 75% 차광처리구 괴근 수는 각각 평균 0개와 0.2개로 감소하였으며, 주당 평근 괴근중도 각각 0 g과 2.7 g으로 괴근 형성이 저해되었다. 지상부 및 지하부 건물중 역시 두 품종 모두 대조구에 비하여 차광 처리 시 감소하였다. 진율미의 경우 지상부 건물중이 대조구에 비하여 55%와 75%차광처리 시 각각 1.2배와 2배 감소하였으며, 지하부 건물중은 각각 5.5배와 6.5배씩 더욱 현저히 감소하였다. 호감미의 경우도 비슷한 경향으로 지상부와 지하부 건물중이 차광처리에 의하여 감소하였다. 반면에 T/R율은 두 품종 모두 차광처리에 따라 증가하는 경향을 보였다.
Table 1.
Effects of shading on the growth pattern of sweetpotato during the storage root formation stage. Sweetpotato plants were grown under different shading treatments (0%, 55%, and 75%) and growth patterns were measured on the 50th day after commencing shading treatments.

괴근비대기(정식 후 50~90일 사이)동안 차광처리 직 후 생육 90일에 수확하여(Fig. 4) 괴근 수량을 조사한 결과(Table 2) 괴근 개수는 진율미 품종의 경우 20 g 이상 괴근 수 및 평균 무게가 각각 6.4개, 61.4 g임에 반해 차광처리구 3.4~ 4.9개, 45.3~51.8 g으로 차광처리에 따라 괴근 비대가 억제되는 경향을 보였고 호감미 품종 역시 비슷한 경향을 보였다. 또한 두 품종 모두 지하부 건물중이 차광처리가 높을수록 감소폭이 크게 나타났다. 그러나 지상부 건물중과 엽면적은 호감미 품종만 75% 차광처리 시에 대조구에 비하여 1.9배 감소하였다. T/R율은 생육초기와 마찬가지로 차광처리에 의하여 두 품종 모두 증가하는 경향을 나타내었다. 이는 차광처리로 인한 낮은 광 조건으로 인해 전체적으로 건물중이 줄어들었을 뿐만 아니라 상대적으로 지상부에 광합성 산물을 더 많이 분배하여 뿌리로 이동되는 광합성 산물이 줄어들어 T/R율은 증가된 것으로 생각되며, 이로 인해 차광수준이 높아질수록 건물중이 낮아진 것으로 판단된다(Kim, 2000; Lee et al., 2019; Song et al., 2014).
Table 2.
Effects of shading on the growth patterns of sweetpotato during the storage root swelling stage. Sweetpotato plants were grown under different shading treatments (0%, 55%, and 75%) during the storage root swelling stage (50–90th day after planting) and growth patterns were determined.
생육시기별 차광 정도에 따른 수확 후 괴근 수량 및 괴근 당도
두 품종 모두 차광처리에 따라 괴근 비대가 저조한 양상을 띠었으며, 특히 호감미가 수량감소폭이 진율미에 비해 상대적으로 크게 나타났다. 또한 차광처리가 높을수록 수확량도 크게 감소되었는데, 55%차광처리에서는 진율미는 5~16%의 수량 감소를 보였으며, 호감미는 15~25%의 수량감소를 나타낸 반면에 75%차광 처리구에서 진율미는 25~39%의 수량감소를 나타냈으며 호감미는 25~52%의 현저한 수량감소가 나타났다. 생육 기간 별로 차광처리에 따라 수량이 감소되는 양상은 차이가 있었다. 괴근 형성기와 괴근 비대성기에 에 비하여 괴근비대기에 두 품종 모두 수량 감소의 큰 영향을 받았다. 괴근비대기 55%와 75% 차광처리구에서 진율미의 경우 대조구 대비 16%와 39%, 호감미의 경우 25%와 52%로 가장 큰 폭의 감소를 보여주었다(Table 3). 이와 같이 괴근 비대기에 일조량이 부족하였을 때 특히 75% 차광처리 시 눈에 띄게 수량이 감소하는 경향을 보였으며 진율미보다 호감미의 수량감소가 더 크게 나타났다. 한편 생육시기별 차광정도에 따른 수확 직후 각 처리별 괴근의 당도를 분석한 결과 진율미 품종의 경우 차광 처리구 괴근당도가 소폭으로 높은 경향을 보였지만 두 품종 모두 유의적인 차이를 보이지는 않았다(Table 4).
Table 3.
Effects of shading treatments on the productivity of sweetpotato. Sweetpotato plants were grown under different shading treatments (0%, 55%, and 75%) and productivity indices were determined at different storage root growth stages.
Table 4.
Effect of shading treatments on the soluble sugar contents in sweetpotato. Sweetpotato plants were grown under different shading treatments (0%, 55% and 75%) and soluble sugar contents were determined at different storage root growth stages.
적 요
최근 기상이변으로 인한 일조부족이 문제되어 작물의 수량 감소 피해가 발생하고 있다. 본 연구는 생육시기에 따라 차광량을 달리하여 고구마의 생육양상 및 수량 감소 정도를 구명하고자 수행하였다.
1. 생육시기별 차광률은 55% 차광 처리시 약 51%, 그리고 75% 차광 처리시에는 약 65% 수준의 차광률을 보였다. 차광처리에 의한 광계 II 암적응 최대양자수율(Fv/Fm) 조사 결과는 호감미와 진율미 두 품종 모두 대조구와 55% 차광 시험구는 유의적인 차이를 보이지 않았고 75% 차광 시험구의 경우 대조구 보다 약간 높은 수치를 보였다.
2. 차광 처리 50일 경과 후 처리별 광합성에 이용되는 색소의 함량을 조사하였다. 호감미와 진율미 두 품종 모두 차광처리구의 엽록소 a와 b의 함량이 대조구에 비하여 유의적으로 증가하는 경향을 나타내었다. 특히 엽록소 b의 함량은 더 많은 비율로 증가 하였다. 호감미의 경우 대조구에 비하여 55%와 75% 차광처리구에서 각각 47%, 41% 증가하였고 진율미의 경우는 각각 39%, 34% 엽록소함량이 증가하였다.
3. 괴근형성기(정식 후 50일간)와 생육 중기(정식 후 50~90일 사이)에 차광처리 시 생육특성을 조사하였다. 지상부 및 지하부 건물중 역시 두 품종 모두 대조구에 비하여 차광 처리 시 감소하였다. 특히 괴근 형성기에 두 품종 모두 수량이 감소하였다. T/R율은 차광처리에 의하여 두 품종 모두 증가하는 경향을 나타내었다. 괴근형성기 및 비대기에 일조량이 부족할 경우 두 품종 모두 전반적으로 괴근 형성 및 비대가 억제되는 경향을 보였다.
4. 수확 후 수량 조사 결과 두 품종 모두 생육시기별 차광처리에 따라 괴근 비대가 저조한 양상을 띠었으며, 특히 호감미가 수량감소폭이 진율미에 비해 상대적으로 크게 나타났다. 또한 차광처리가 높을수록 수확량도 크게 감소되었다. 괴근 형성기와 괴근 비대성기에 비하여 괴근비대기에 두 품종 모두 수량 감소의 큰 영향을 받았다.
