

옥수수(Zea mays L.)는 일반적으로 생육기간 동안 따뜻한 온 도 및 기후 조건을 필요로 하는 식량작물이다. 옥수수는 식량과 사료용 외에도 각종 산업 및 공업용, 특히 바이오 에너지의 원 료로서 많이 이용되고 있다(Chen et al., 2013; Vohra et al., 2014). 하지만 최근 지구온난화와 슈퍼 엘리뇨로 인한 기온 상 승 및 급격한 기후 변화로 작물 생육에 재해 및 피해가 지구촌 곳곳에서 발생되고 있으며, 최근 지속되는 가뭄에 우리나라에 서도 논 물 마름 현상, 작물 시듦, 생육저조 현상 등의 피해가 발생하고 있다. 기상이변 중 폭염과 가뭄은 심각한 한발 피해로 이어져 특히 옥수수의 생육을 저해하며, 수량 또한 현저히 감소 시킨다.
한발은 장기간에 심한 강수량 부족으로 인하여 발생하는 기 상재해로서, 비가 적게 내리거나 강한 햇볕이 계속되면 물의 균형이 깨지면서 물 부족 현상이 나타난다. 특히 옥수수는 키 가 크고 잎이 넓은 작물로서 유묘기에 극심한 한발 스트레스를 받으면 잎말림 현상이 나타나 생육이 저조하며 (Song et al., 2015), 생육기간 중 개화기 전후로 물 부족 현상은 수량에 큰 영향을 미치므로 가뭄이 들면 물을 대주어야 한다. 또한 수정 후 등숙 기간에 한발피해가 발생하면 옥수수 종실이 작고 수량 이 현저히 감소하여 상품가치 하락으로 출하가 어렵게 된다. 하지만 토양이 과습 상태로 오랫동안 유지되면 습해를 받아 내 부의 산소부족이 발생하여 뿌리가 썩게 될 수 있으므로, 장마 철에는 배수관리에 신경을 써야 한다.
옥수수는 수분 부족으로 한발 스트레스를 받으면 유묘기 형 성, 초기영양생장, 광합성능력, 생식생장, 유수형성, 수정, 종실 형성 및 수량 등에 상당한 영향을 미치게 된다(Aslam et al., 2013). 옥수수 잎 발달이 저해되면 잎 면적이 감소하여 광합성 효율이 떨어지며, 유수형성기 이후에 한발 스트레스를 받으면 암이삭이 수꽃보다 늦게 발달하여 수분이 어려워 불임이삭이 발생한다. 또한 한발 스트레스로 인한 고온현상은 수꽃의 개화 유도를 방해하며, 꽃가루 발달을 저해하는 요인으로 나타날 수 있다(Herrero and Johnson, 1981).
일반적으로 옥수수 한발 내성에 관한 주요 관심은 anthesissilking interval (ASI)과 종실 성숙에 집중되고 있으며, 특히 ASI는 전통적으로 한발과 연관된 지표로서 옥수수 연구자들에 의해 사용되었다(Bolaños and Edmeades, 1996). ASI는 주로 수꽃과 암꽃의 개화기 차이를 나타내며, 특히 한발 스트레스 환경에서는 수꽃과 암꽃의 개화기가 정상환경 보다는 차이가 벌어지는 것으로 나타난다(NeSmith and Ritchie, 1992). 이러 한 한발 스트레스를 줄이기 위해서 관개가 가장 효과적인 방법 이지만, 한정된 물 자원과 비용 등에 대한 문제로 가뭄에 대한 충분한 관개가 쉽지 않다. 토양 수분 부족은 영양생장기와 수 량관계에 상당히 영향을 미치는 것으로 나타났으며, 특히 수꽃 의 발달 시기와 옥수수 형성 단계에서 수분 부족은 건물중 감 소와 수량감소로 나타났다(Cakir, 2004). 최근에는 한발 피해 를 줄이기 위해서 다양한 생장조절제를 이용한 처리 효과 (Saruhan et al., 2012; Zamaninejad et al., 2013; Seo et al., 2014), 고온 스트레스에서 포스파티드산(phosphatidic acid)의 발현양 연구(Chen et al., 2010), 한발 저항성 및 감수성을 가 진 옥수수에서 한발 처리를 통한 옥수수잎 대사산물 비교 및 한발 반응연구(Barnaby et al., 2013) 등이 있다.
한발에 대한 내성은 다양한 유전자가 연관되는 복잡한 특징 을 가지고 있으며, 이러한 유전자원의 다양성과 계통 간의 유 연관계는 유전체의 발달로 인하여 신품종 개발에 이용될 수 있 다(Lee and Tracy, 2009). 유전체 기술은 한발 저항성 옥수수 육종과 선발에 이용할 수 있으며, 또한 분자 마커 및 형질전환 기술을 작물 육종에 적용하여 원하는 개체 선발이 가능하게 되 었다(Cattivelli et al., 2008; Langridge and Reynolds, 2015).
과거에는 표현형을 이용한 표지 인자를 통하여 선발하였으나, 현재는 DNA를 이용한 분자 마커 시스템이 활성화되면서 정밀 한 유전자 지도와 분자 마커가 유전적 연관 등에 대한 연구가 활발하게 진행되고 있다.
본 논문에서는 가뭄 및 수분 부족으로 인하여 발생되는 옥수 수 한발 관련 연구에 대하여 알아보고, 또한 한발에 대한 생리 적 및 분자적 접근분석과 유전체 기술을 활용한 옥수수 연구 현황에 대한 정보를 제공하고자 한다.
한발이 옥수수 생육에 미치는 영향
옥수수는 다른 작물들과 달리 전 생육기간 동안 비교적 많은 양의 물을 필요로 하며, 토양수분이 부족할 경우에는 생육저조 및 수량에 큰 영향을 미친다. 옥수수는 생육초기에 극심한 수 분 스트레스를 받으면 잎말림이 진행되어 생장이 지연되거나 억제되며, 수량감소의 원인이 되기도 한다. 특히 수꽃이 나오는 시기(tasseling)와 수정(pollination)이후 유숙기(milk stage)에 수분 스트레스를 받으면 잎 시듦 및 말림이 증가되고, 도복이 증가하여 수량 저조로 이어진다(Table 1, Rhoads and Bennett, 1990; Shaw, 1988).
Table 1.
Estimated corn evapotranspiration and yield loss per stress day during various stages of growth.
영양생장 생육단계 중에서 V8~V16 (잎이 모두 나오고, tasseling 이 시작하는 단계: 식물 발생 후 4주에서 66일 사이)은 이삭 사이즈(ear size)와 알갱이 수를 결정하는 단계이다. 특히 이삭 사이즈는 V8~V14 단계에서 결정되는데, 이 시기에 한발 스트 레스를 받으면 이삭 사이즈와 수량이 감소하게 되며, 잠재적으 로 수량은 10~30% 정도 줄어들 수 있다(Heiniger, 2000). 그 리고 V14~tasseling 단계는 수정이 될 수 있는 알갱이 수를 결 정하는 시기이며, 이 때 한발 스트레스를 받으면 옥수수 수량은 최소 10%, 최대 50%까지 감소할 수 있다(Fig. 1, Heiniger, 2000). Fig. 1은 영양생장 V8~V16 (tasseling) 시기 동안에 수 분 스트레스(한발 기간)와 잠재적 옥수수 수량 손실의 관계를 보여준다. Tasseling 단계는 한발에 민감한 시기이며(Westgate and Grant, 1989), tasseling 이전에 수분 스트레스를 받으면 tasseling과 silking의 발생이 2주 이상 지연되고, 90% 이상의 수량이 감소된다(NeSmith and Ritchie, 1992).
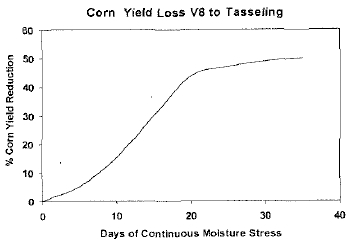
Fig. 1.
Corn yield loss due to moisture stress from V8 to tasseling. Figure based on Heiniger (2000).
Udomprasert et al. (2005)은 tasseling 초기단계에 수분 스트 레스를 받으면 ASI가 2~3일 증가하였고, 반면 개화기(anthesis) 단계에 수분이 부족하면 ASI는 0.75~1.25일 증가하였다. 결론 적으로 수분이 부족하면 ASI는 증가하고, 이삭 둘레, silk 성 장, 종자 무게, 곡실 수량, 수확 지수 등은 감소하는 것으로 나 타났다.
생식생장 기간에 한발 스트레스로 인한 수량 손실을 살펴 보 면, 개화 및 수정 단계에서 피해를 받을 때 출사기(silking)가 늦고, 옥수수 수염 길이가 줄어들고, 수정 후 배 발생이 억제된 다(Westgate and Boyer, 1985; Lu et al., 2011). 특히 수정을 하는 이 시기에 수분 스트레스를 받으면 화분(꽃가루) 방출과 옥수수 수염의 동시 발생이 억제 및 방해를 받아 옥수수 수량 은 3~8% 감소한다(Table 1). 또한 등숙 기간에 한발 스트레스 는 낟알 무게에 영향을 미친다. 이것은 수분 스트레스로 인하 여 잎이 시들거나 죽어가고, 등숙 기간이 짧아지고, 도복이 증 가하여 수량이 감소하기 때문이다. 결국 등숙기 동안에 수분 스트레스를 받으면 하루 수량 감소율이 3.0~5.8% 감소하며, 이 기간에 극심한 수분 스트레스를 받으면 옥수수 수량은 20~30%까지 감소 할 수 있다.
또한 옥수수 뿌리는 여러 가지 환경에서 중요한 구성요소로 작용하고 있는데, 식물이 물과 미네랄 등을 흡수할 수 있도록 도와 주며, 각 생장 및 생육 단계 중에서 유묘기의 뿌리는 한발 내성과 중요한 요소로 작용한다(Weerathaworn et al., 1992). Morizet et al. (1983)은 수분 스트레스가 옥수수 뿌리 생장에 영향을 미치며, 특히 뿌리의 무게가 잎의 무게보다 더 증가하 였음을 알아냈다. 뿐만 아니라 옥수수의 영양생장기와 뿌리 생 장 단계에서 한발 스트레스는 잎과 뿌리의 생장을 감소시키는 것으로 확인하였다(Ramadan et al., 1985).
한발 관련 생리적 및 분자적 연구
Aslam et al. (2006)은 옥수수의 발아와 생육초기 단계에서 수분 스트레스의 저항성 평가를 위하여 relative cell membrane injury (RCI %age)와 stomatal conductance를 측정하였으며, 이들의 두 가지 지표는 내건성을 위한 선발기준으로 활용할 수 있는 것을 알아냈다. 한발에 영향을 주는 생리적으로 관련된 요 인은 식물 조직의 수분 함량과 수분 포텐셜이다(Jones, 2007). Table 2는 수분 부족 및 한발 조건에 관련된 생리적 특성들을 다양한 범위로 보여주고 있으며, 모든 한발 환경조건에서는 수 량과 매우 연관되어 있는 것을 알 수 있다(Cattivelli et al., 2008).
Table 2.
Physiological traits relevant for response to drought conditions.
| Plant traits | Effects relevant for yield | Modulation under stress | References |
|---|---|---|---|
| Stomatal conductance/ leaf temperature | More/less rapid water consumption. Leaf temperature reflects the evaporation and hence is a function of stomatal conductance | Stomatal resistance increases under stress | Jones 1999, Lawlor and Cornic 2002 |
| Photosynthetic capacity | Modulation of concentration of Calvin cycle enzymes and elements of the light reactions | Reduction under stress | Lawlor and Cornic 2002 |
| Timing of phenological phases | Early/late flowering. Maturity and growth duration, synchrony of silk emergence and anthesis, reduced grain number | Wheat and barley advanced flowering, rice delayed, maize asynchrony | Slafer et al., 2005, Richards 2006 |
| Anthesis-silking interval (ASI) in maize | ASI is negatively associated with yield in drought conditions | Drought stress at flowering causes a delay in silk emergence relative to anthesis | Bola os and Edmeades 1993, Edmeades et al., 2000 |
| Starch availability during ovary/embryo development | A reduced starch availability leads to abortion, reduced grain number | Inhibition of photosynthetic activity reduces starch availability | Boyer and Westgate 2004 |
| Partitioning and stem reserve utilization | Lower/higher remobilization of reserves from stems for grain-filling, effecting kernel weight | Compensation of reduced current leaf photosynthesis by increased remobilization | Blum 1988, Slafer et al., 2005 |
| Stay green | Delayed senescence | Rajcan and Tollenaar 1999 | |
| Single plant leaf area | Plant size and related productivity | Reduced under stress (wilting, senescence, abscission) | Walter and Shurr 2005 |
| Rooting depth | Higher/lower tapping of soil water resources | Reduced total mass but increased root/shoot ratio, growth into wet soil layers, regrowth on stress release | Hoad et al., 2001, Sharp et al., 2004 |
| Cuticular resistance and surface roughness | Higher or lower water loss, modification of boundary layer and reflectance | Kerstiens 1996 | |
| Photosynthetic pathway | C3/C4/CAM, higher WUE and greater heat tolerance of C4 and CAM | Cushman 2001 | |
| Osmotic adjustment | Accumulation of solutes: ions, sugars, poly-sugars, amino acids, glycinebetaine | Slow response to water potential | Serraj and Sinclair 2002 |
| Membrane composition | Increased membrane stability and changes in aquaporine function | Regulation in response to water potential changes | Tyerman et al., 2002 |
| Antioxidative defense | Protection against active oxygen species | Acclimation of defence systems | Reddy et al., 2004 |
| Accumulation of stressrelated proteins | Involved in the protection of cellular structure and protein activities | Accumulated under stress | Ramanjulu and Bartels 2002Cattivelli et al ., 2002 |
수분부족은 광합성에 영향을 미쳐 식물생육과 수량에 영향 을 주며(Schussler and Westgate, 1991), 특히유묘기 단계에 서는 시들음, 발육 방해, 출엽 지연, 잎면적 감소 등의 원인이 된다(Abrecht and Carberry, 1993). 옥수수는 개화기 화분방 출과 옥수수 수염 발생이 서로 밀접하게 연관되어 있기 때문에 이 시기에 수량의 정도가 결정되어지며, 최종 수량과 ASI 간격 의 정도 사이에는 긴밀한 관계가 성립된다. 따라서 개화기에 한발 스트레스는 옥수수 수염의 발생을 늦춰주는 원인이 된다 (Bolaños and Edmeades, 1993; Ribaut et al., 1996). 많은 연구자들이 한발 내성 품종을 확인하기 위하여 CMMYT에서 제시된 방법을 이용하여 선발지표로 활용하고 있다. Bänziger et al. (2000)은 옥수수 한발 내성 선발을 효율적으로 돕기 위 하여 다양한 이차 형질들을 제시하였다(Table 3).
Table 3.
Secondary traits to identify drought tolerance.
Obeng-Bio et al. (2011)은 잎 상대 수분함량, 잎말림, 잎노 화, ASI, 옥수수 이삭수, 수량 등의 이차 형질을 이용하여 한발 내성 옥수수 품종의 선발을 위한 효과적인 지표를 확인하였다. Song et al. (2015)은 옥수수 유묘기에서 수분 스트레스에 따 른 잎말림을 측정하여 한발 내성 평가 및 선발을 위한 지표를 확인하였으며, 결과적으로 두 번째와 세 번째 잎의 잎말림 평 균이 2.5 이하를 보이는 품종은 유묘기에 한발 내성이 강한 것 으로 보였다. 이 밖에 한발 내성에 대한 간접적인 선발 지표로 서, antioxidant system (Li et al., 1998; Chugh et al., 2011), salicylic acid / abscisic (Saruhan et al., 2012; Seo et al., 2014), polyethylene glycol (PEG) (Khodarahmpour, 2011), oxidase oxalate activity / root elongation (Voothuluru et al., 2013), phosphatidic acid (Chen et al., 2010), potassium (Aslam et al., 2013) 등을 이용하여 다양한 선발 지표들을 제 시하고 있다. 최근에는 옥수수 뿌리에 한발 스트레스를 받을 때 phytoalexin 물질이 유발되며, 이 물질은 한발과 관련 있다 고 보고하였다(Vaughan et al., 2015). Phytoalexin은 외부 독 성에 대해 식물조직이 생산하는 독성 저항물질 및 항생물질로 서, 옥수수 뿌리에서 한발상태가 계속 유지되는 동안 zealexins 와 kauralexins 물질의 양이 계속해서 증가하였으며, 이 물질 들의 축적은 한발 내성과 관련 있는 것으로 밝혀냈다. 이러한 지표들은 이차 특성들과 비교했을 때, 연구자들이 많은 시간과 노동력이 필요하지 않아 한발 내성 평가에 많이 활용되고 있는 추세다.
유전체를 활용한 옥수수 한발 내성 연구
1980년대에는 DNA를 기본으로 한 분자 마커가 옥수수 육 종을 향상시킬 수 있는 가능성을 확인하였으며(Helentjaris et al., 1985; Paterson et al., 1988), Botstein et al. (1980)에 의 해 restriction fragment length polymorphisms (RFLPs)이 확 인되어 일반적으로 분자 육종이라는 새로운 연구 분야를 발달 시켰으며, 양적 형질을 위한 유전적 획득을 향상시키는데 이용하 였다. 1990년대에는 양적 형질의 향상을 위해 marker assisted selection (MAS)을 이용한 연구가 시작되었으며, 이것은 표현 형 및 유전형 정보를 합친 선발 방법으로서 엘리트 육종 집단 에서 평균 성능을 증가시켰다(Lande and Thompson, 1990). 또한 Marker assisted breeding 프로그램은 전통적인 육종 프 로그램과 비교하여 데이터와 분석의 양이 약 7배나 증가하였으 며, 육종가들은 marker assisted recurrent selection (MARS) 을 이용하여 일년에 한 두 번 실시했던 기존의 육종 방법보다 세네 번 정도 육종 선발을 결정할 수 있게 되었다(Eathington et al., 2007). 이렇게 발전한 유전적 개량으로는 recombinant DNA technology, genomics, bioinformatics 등이 있으며, 이 것을 이용하여 작물의 품종 선별 및 육종에 다양하게 이용되고 있다.
한발 스트레스는 옥수수 육종 및 생산에 큰 영향을 미치는 스트레스 조건이다. 또한 기후온난화와 강수량이 불규칙해지면 서 전세계는 점차 한발에 견딜 수 있는 옥수수가 필요하게 되 었다. 특히 영양 생장 기간에는 수분 스트레스에 민감한데, 이 시기에 수량이 결정되기 때문이다. 이와 같이 수량 관련 형질 로 옥수수 QTL 연구가 시작 되었으며, 농업적으로 필요한 다 양한 형질에 대한 분자 마커를 개발하였다(Stuber et al., 1987). 분자 마커는 수량, 환경 스트레스, 병충해, 고온 및 한 발 내성 등 다양한 형질에 연관된 유전적 정보를 관찰할 수 있 으며, 특히 개체들 사이의 유전적 연관성을 추정할 수 있다 (maizeGDB, http://www.maizegdb.org/assembly/). 또한 QTL 의 유전적 비교분석을 통하여 특정 형질에 연관된 유전자의 수, 염색체 상의 위치, 상위작용, 그리고 각 유전자의 영향력 등 의 다양한 정보를 GRAMENE 웹사이트(http://www/gramene.org/qtl/)에서 확인 할 수 있다. QTL 분석을 이용한 한발 내성 에 대한 유전적 분석은 1995년에 시작되었으며(Lebreton et al., 1995), 이후 분자 마커를 이용하여 스트레스와 비 스트레 스에 관여하는 유전체를 발굴하여 연구 목적에 맞는 집단을 찾 을 수 있게 되었다. 또한 옥수수의 비 생물학적 환경스트레스 내성과 관련된 유전자 및 QTL 데이터를 Plantstress 웹사이트 (http://www.plantstress.com/biotech/index.asp?Flag=1)를 통 하여 검색할 수 있으며, 이 사이트에서 수집된 데이터를 기반 으로 하여 옥수수 한발 스트레스에 연관된 QTL 데이터 및 자 료를 Table 4에서 확인할 수 있다.
Table 4.
Summary of QTLs identified for drought tolerance-related traits in maize.
| Specific root and shoot responses under drought | ||||
|---|---|---|---|---|
| Trait | Population | Type | QTL | Reference |
| Leaf elongation rate in correspondence with ASI | Ac7643 and Ac7729/TZSR | RIL | 5 | Welcker et al., 2007 |
| Leaf elongation under stress in several populations Several | 19 | Welcker et al., 2011 | ||
| Leaf temperature and seedling DM under drought stress | Zong3 x 87-1 | RIL | 9 | Liu et al., 2011 |
| Roots traits and yield | Lo964 x Lo1016 | F3 | <11 | Tuberosa et al., 2002 |
| Roots traits and yield | Lo964 x Lo1016 | NIL | 1 | Landi et al., 2010 |
| Various root traits | 3 populations | RIL | 15 | Burton et al., 2014 |
| Seedling root traits in PEG solution | Ac7643 x Ac7729/TZSRW | RIL | 13 | Ruta et al., 2010 |
| Seedling survival | BC inbred lines of 11 crosses | BC2F2 | 7 | Hao et al., 2009 |
| Various shoot & root traits | Tropical population | RIL | 30 | Messmer et al., 2011 |
| Whole plant and crop drought resistance | ||||
| Adaptive and constitutive traits | 12 populatios x 22 locations | Many | Hao et al., 2010 | |
| Flowering parameters under drought, ASI | Ac7643S5 X Ac7729/TZSRWS5 | F2 | 7 | Ribaut et al., 1996 |
| Plant height, ear number, days to silking, yield | SD34 X SD35 | F3 | 5 | Agrama and Moussa 1996 |
| QTL × environment interaction | F2-F3- | Vargas et al., 2006 | ||
| Yield and ASI under field drought | X178 × B73 | F2-F3- | 2 clusters (45 & 60 QTLS) | Hao et al., 2008 |
| Grain yield and ASI over locations | 3 populations | 83 & 62 | Almeida et al., 2013 | |
| Grain yield and components | 350 crosses | Many | Xue et al., 2013 | |
| Hormonal responses under drought | ||||
| Leaf ABA concentration | Os420 X IABO78 | F3 | 16 | Tuberosa et al., 1998 |
| Leaf ABA concentration, yield | Os420 X IABO78 | F4 | 17 | Sanguineti et al., 1999 |
| Water-use and photosynthetic activity under drought | ||||
| Adventitious root formation under waterlogging | B64 × teosinte | F2 | 2 | Mano et al., 2005 |
| Vacuolar invertase activity | F2 X Io | RIL | 18 | Pelleschi et al., 1999 |
QTL은 양적 형질에 연관되어 개념적으로 한 개의 형질에 영향을 미치는 연관(linkage)된 유전자 집단이다. Sari-Gorla et al. (1999)은 옥수수 한발 내성 평가를 위하여 male flowering time (MFT), female flowering time (FFT), ASI, plant height (PH) 및 분자 마커 사이에서의 유전적 연관을 분석하였으며, MFT와 PH에서는 대부분의 QTL이 같은 염색체 상에서 발견 되었다. 그에 반해 FFT와 ASI는 한발 스트레스뿐만 아니라 정 상 조건에서도 다양한 QTL이 연관지도에서 발견되었다. Ribaut et al. (1996)은 열대 옥수수를 이용하여 한발 상태에서 flowering parameter와 ASI의 관계를 QTL로 확인하였으며, 수분 스트 레스 조건에서 male flowering (MFLW)와 female flowering (FFLW)은 4개의 QTL, ASI와 MFLW은 1개의 QTL, 그리고 ASI와 FFLW는 4개의 QTL이 공통적으로 나타났다. 한편 한 발 조건에서 잘 견디는 옥수수를 개발하기 위하여 ASI 기간을 단축 시키는 것은 효율적인 육종 선발 방법이다(Xiao et al., 2004). 이 연구팀은 한발 스트레스를 받은 옥수수에서 flowering time QTL을 확인하기 위하여 simple sequence repeat (SSR) 마커를 이용하였으며, 결론적으로 한발 상태에서 MET는 9개, FFT는 6개, 그리고 ASI는 6개의 QTL이 확인되었다. 또한 한 발 조건에서 한발 내성과 수량요소 관계를 QTL로 분석을 한 결과, 곡실 수량은 1개, 백립중은 5개, 이삭(ear) 당 낟알 수는 2개, 이삭당 옥수수속(cob) 무게는 6개, 이삭당 낟알 무게는 1 개, 이삭무게는 3개, 한 개체당 이삭수는 2개 등의 QTL이 발 견되었다(Xiao et al., 2005). Gemenet et al.(2010)은 한발 감수성 품종(KCB)과 한발 내성 품종(GBK032357)을 교배하 여 203개의 F2 집단을 선발하였으며, F2 집단의 short ASI(한 발 내성)와 long ASI (한발 감수성)를 확인하기 위하여 bulked segregant analysis (BSA)으로부터 ASI와 연관된 SSR 마커를 선발하였다. 결과적으로 ASI에 관련된 4개의 마커는 p-umc2189, p-umc1542, p-bnlg1179, p-bnlg1014 등을 확인하였으며, QTL 분석을 통하여 p-umc2189와 p-bnlg1014는 FFT와 연관이 있 으며, p-umc1542와 p-bnlg1179는 kernel number (KN)와 grain yield (GY)에 연관되어 있음을 확인하였다. 또한 한발 내성 형질을 가진 이 마커들은 엘리트 품종에 적용할 수 있는 MAS 개발에 유용하게 이용할 수 있다고 보고하였다.
과거에는 수분 부족 상태(수분 스트레스)에서 옥수수 잎과 뿌리의 전사 변화를 설명하기 위해서 microarray chip hybridization 방법을 주로 사용하였으며(Zheng et al., 2004; Poroyko et al., 2007; Spollen et al., 2008), 최근에는 next-generation sequencing이 발달하여 RNA sequencing (RNA-Seq)을 이용 한 양적 유전자의 발현을 분석하고 있다(Metzker, 2010). Opitz et al. (2014)은 low water potential에서 유묘기 옥수수 주근 의 transcriptional response (전사반응)를 RNA-Seq을 이용하 여 확인하였다. 단기 수분 스트레스(6 h)에서 약한 수분 부족 (-0.2 MPa)과 극심한 수분 부족(-0.8 MPa)은 각각 249와 3,000개의 유전자가 발현하였으며, 장기 수분 스트레스(24 h) 에서는 7,267(-0.2 MPa)과 12,838(-0.8 Mpa)개의 유전자가 단기 수분 스트레스에서 반응한 유전자의 80% 이상을 포함하 였다. 한편 한발 내성을 위한 옥수수 phenotyping은 비용이 들 거나 시간이 많이 소요된다. Ziyomo and Bernardo (2013)는 한발 조건에 있는 옥수수에서 수량과 이차 특성을 위하여 유전 력, 유전분산, 유전상관 등을 밝혀냈고, 옥수수 한발 내성에 대 하여 이차 특성과 Genomewide selection을 통하여 간접적인 선발의 효율성을 알아냈다. 결과적으로 한발 내성 선발을 위하 여 phenotyping 보다는 마커를 기초로 한 genotyping이 효율 적이며 비용도 감소하였다. 또한 genomewide selection은 1년 에 3세대까지 최대로 수행할 수 있게 되었다(Bernardo and Yu, 2007; Eathington et al., 2007; Bernardo, 2008). 한편, 몬산 토에서는 유전자 조작 기술을 이용하여 가뭄 저항성의 특성을 가진 옥수수 제품인 Genuity® DroughtGard™을 개발 및 생 산하고 있으며, 단 2개의 가뭄 내성을 지닌 GM 작물이 허가 되었고, 상업적으로 생산하고 있다(Langridge and Reynolds, 2015, Table 5).
Table 5.
Drought tolerance crops approved for cultivation.
Gulli et al.(2015)은 GM 옥수수(DKC6575, Monsanto)와 near-isogenic non-GM 옥수수(cv. Tietar)에서 한발 스트레스 반응과 유전자 발현을 비교하였으며, 한발 스트레스를 받은 두 품종 모두 순광합성량, 기공전도, 엽면증산 등 크게 감소하였 다. 또한 한발 스트레스에서 229개의 유전자가 up-regulated 하였으며, Tietar 품종에서 170개, DKC6575 품종에서 9개, 그리고 두 품종 모두 공통적으로 50개의 유전자가 발현되었다.
적 요
한발은 환경 스트레스에 미치는 자연재해로서, 장기간에 걸 쳐 강수량 및 수분공급이 저하되면 수분이 결핍되어 작물이 정 상적인 생리활동을 할 수 없고, 생육 또한 불량하여 수량감소 에 큰 영향을 미친다. 모든 작물들이 한발의 영향을 크게 받지 만 어느 생육 단계에서 한발 스트레스를 받느냐에 따라서 피해 가 달라진다. 일반적으로 옥수수는 한발 스트레스를 받으면 유 묘기 형성, 영양 생장, 뿌리 발달, 광합성, 개화기, ASI, 종실 형성, 수량 등에 심각한 영향을 미치게 된다. 특히 영양생장에 서 생식생장으로 전환되는 단계에 한발 스트헤스를 받으면 수 꽃 및 화분 방출이 늦어지고, 출사기 및 옥수수 수염 발생도 늦 어져 ASI가 증가하여 수정이 불가능 하거나 수정이 되더라도 배 발생 억제 및 방해를 받아 수량 감소의 큰 원인이 된다. 이 러한 한발에 대한 피해를 줄이기 위해서 1980년대부터 최근까 지 마커와 표현형이 연관된 유전체를 바탕으로 다량의 분자적 데이터 분석을 통한 옥수수 한발 내성 품종 선별 및 육종에 대 한 많은 연구들이 진행되고 있다. 또한 최근에는 수량 등 다양 한 유전자들이 관여하고 환경 스트레스에 영향을 받는 양적형 질 QTL에 관한 많은 연구들이 수행되고 있으며, genomics 분 야에서 신기술인 MAS를 이용하여 목표유전자 이입 및 선발을 통해 또 다른 육종 선발 도구로 활용되고 있다. 뿐만 아니라 유 전자 조작기술을 이용한 한발 내성 특징을 가진 옥수수를 개발 하여 제품으로 생산 및 판매되고 있다. 과거 전통적인 육종 방 법은 자식계통의 표현형 분석과 특별 조합을 통한 계통으로부 터 데이터를 분석하여 품종을 육성하였으나, 현재는 모든 작물 의 유전체 전체를 이용한 데이터 베이스와 분자 마커 기술을 한발 내성 육종에 활용하여 다양한 연구가 이루어지고 있다. 이러한 분자적 육종 기술의 발달은 우수한 연구 결과를 도출 및 확보할 수 있으며, 옥수수 한발 내성 신품종 개발에 있어서 새로운 육종 기술로 적용할 수 있을 것이다.
