

Due to climatic changes caused by global warming, high temperature and substantial precipitation will directly impact crops and pests in agriculture (Chakraborty et al., 2000; Morton, 2007; Legzdina et al., 2013). Untimely rainfall occurring during the rice ripening stage induces serious problems. One of the most significant problems is pre-harvest sprouting, the germination of mature seeds while still on the mother plant, which occurs in wet or humid conditions prior to harvest (Rodríguez et al., 2009; Chono et al., 2013; Liu, 2013). These also influence the pre-harvest sprouting of seeds, which in turn affect the quality and production of rice.
Temperature and moisture are the main environmental factors affecting pre-harvest sprouting. The major factors affecting pre-harvest sprouting tolerance other than environmental conditions are seed dormancy, seed coat permeability and color, α-amylase activities, and endogenous hormones levels (Chen et al., 2008; Gao et al., 2013).
Seed dormancy at harvest is a desired trait because it prevents the early germination of grains in the head following exposure to moist conditions. However, excessive dormancy can also be problematic because treatments might be required to promote germination (Gubler et al., 2005; Finch-Savage and Leubner- Metzger, 2006; Schenkelaars, 2007). Seeds subject to pre-harvest sprouting offer reduced seed viability and hydrolysis of starch and protein in the endosperm, which creates a favorable environment for serious infection by saprophyte fungi. This causes not only reduced grain yield, but also damages the quality of the end-product, resulting in economic losses (Gubler et al., 2005; Yang et al., 2007; Huang et al., 2012).
The accelerated aging test is recognized as an accurate indicator of seed vigor and storability (Tekrony, 2003; Abdellaoui et al., 2013; Parmoon et al., 2013) and is very effective in testing the relative storage potential of seed lots (McDonough et al., 2004; Babiker et al., 2010). High vigor seed lots withstand accelerated aging stress and deteriorate more slowly than low vigor seed lots. The germination differences of high and low quality seed lots after accelerated aging showed a similar trend to the germination of the same seed lots after warehouse storage. With longer storage periods, the progressive loss of seed quality reduces the rate of germination and eventually results in loss of viability (Thant et al., 2010; Bewley et al., 2013). In the present study, the influence of rainfall during the ripening stage on pre-harvest sprouting and seed viability was investigated, and the seed longevity of rainfall-treated seeds by accelerated aging test was reported in two Korean-bred rice cultivars.
Materials and Methods
Plant materials
Two medium-late maturing japonica rice cultivars, Shindongjin and Hopum, high quality japonica-type rice cultivars, were used in the experiment. Shindongjin is a low-tillering heavy panicle type, whereas Hopum is an active tillering type cultivar. In a former experiment on pre-harvest sprouting of the two varieties, the pre-harvest sprouting resistance of Shindongjin was higher than that of Hopum because the seed dormancy of Shindongjin was much higher than that of Hopum at the ripening stage (Bark and Chung, 2014). The two varieties were cultivated in an experimental field of Chonbuk National University located at the latitude of 35° 49’ and longitude of 127° 9’ in Jeonju-city in 2012. The 30-day nursery seedlings of rice cultivars were transplanted Jun. 1 and were harvested at 65 days after heading (DAH) (Oct. 19). The heading date of both cultivars was Aug. 15.
Artificial rainfall treatment and pre-harvest sprouting survey
The influence of rainfall during the ripening stage on preharvest sprouting was investigated. For the rainfall treatment, rice plants (20 hills a treatment) grown in the field were moved to a side-opened greenhouse, some at 40 DAH (Sept. 25) and some at 50 DAH (Oct. 5). The greenhouse was equipped with vertical spraying sprinklers. Artificial rainfall was applied to the rice plants for different durations of 0, 3, 5, 7 or 10 days. The percent of pre-harvest sprouting was investigated at harvest (64 DAH).
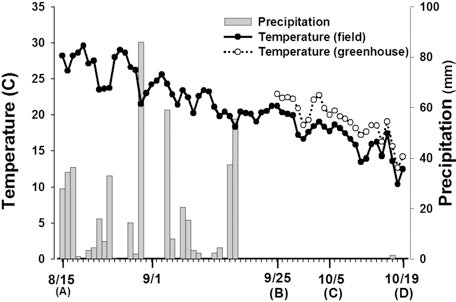
Meteorological data in the field and greenhouse during the experiment period is shown in Fig. 1. The average temperature during the ripening stage was 21.0°C, and the total precipitation was 477.7 mm (Aug. 15 to Oct. 19). Average field and greenhouse temperatures during the treatment period (from Sep. 25 to Oct. 19) were 16.8°C and 19.2°C, respectively. The average greenhouse temperature was about 2.4°C higher than that of the field during the same period.
Seed viability, vigor, and longevity of artificial rainfall-treated seeds
To investigate the quality change in harvested seeds treated with artificial rainfall during the ripening stage, seed germination, vigor, and accelerated aging tests were conducted. The seeds were harvested and dried at room temperature to an approximate 14% moisture content, and then germinability and vigor were tested. Germination testing was conducted at 25°C for 14 days with four replicates of 100 seeds each, and vigor was evaluated by germination percentage at five days after imbibition (ISTA, 2009).
In the accelerated aging test, the same germination test procedures as above were followed, except high temperature and high relative humidity treatments were included before the germination test. The seeds were placed in a desiccator containing 100 ml of distilled water, and the desiccator was kept in an incubator at 40±1°C for 30 days (Kim 2004). Subsequently, the seeds from the desiccator were sampled,
and the seed viabilities were tested at five-day intervals.
Statistical analysis
The experiment was arranged in a completely randomized design (CRD) with four replicates, and all collected data were subjected to analysis of variance using Statistical Analysis System (SAS 9.1). The treatment means were compared using Duncan’s Multiple Range Test (DMRT) or t-test.
Results
Pre-harvest sprouting according to artificial rainfall periods
The influence of rainfall during the ripening stage on preharvest sprouting was investigated, and the results are shown in Table 1. The timing of artificial rainfall treatment was determined at 40 DAH and 50 DAH on the basis of seed germinability after heading. Seed germinabilities were different at 40 DAH (26.3% in Shindongjin and 63.9% in Hopum) but similar at 50 DAH (about 76% in both cultivars), as shown in Table 1. When the artificial rainfall was administered at 40 DAH, Hopum showed 1.8% pre-harvest sprouting at three days of rainfall treatment (DRT) and 7.0%, 15.0%, and 15.8% at five, seven, and ten DRT, respectively. Pre-harvest sprouting increased with an increase in the rainfall period. On the other hand, Shindongjin did not demonstrate pre-harvest sprouting at 40 DAH, regardless of rainfall treatment.
Table 1.
Percentage of pre-harvest sprouting seeds after harvest according to rainfall duration during the ripening stage in rice.
| DAH | Variety | Duration of artificial rainfall treatment (days) | ||||
|---|---|---|---|---|---|---|
| 0 | 3 | 5 | 7 | 10 | ||
| ------------------------------------------------------ % ------------------------------------------------------ | ||||||
| 40 | Hopum | 0.0 A*d** | 1.8 Bc | 7.0 Ab | 15.0 Aa | 15.8 Ba |
| Shindongjin | 0.0 Aa | 0.0 Ca | 0.0 Ba | 0.0 Ba | 0.0 Da | |
| 50 | Hopum | 0.0 Ad | 6.3 Ac | 7.3 Ac | 15.3 Ab | 25.8 Aa |
| Shindongjin | 0.0 Ab | 0.0 Cb | 0.0 Bb | 0.3 Bb | 0.8 Ca | |
At 50 DAH, the pre-harvest sprouting of Hopum was higher than that at 40 DAH, but the germination trends were similar. The pre-harvest sprouting was 6.3% at three DRT, 7.3% at five DRT, 15.3% at seven DRT, and 25.8% at ten DRT. In contrast, pre-harvest sprouting of Shindongjin was only observed in the plants receiving more than seven days of rainfall at 50 DAH, and its germination percentages were also very low (0.3% at seven DRT and 0.8% at ten DRT) compared to those of Hopum.
Seed vigor and germinability after harvest
The vigor and germinability of harvested seeds treated with artificial rainfall at 40 DAH and 50 DAH are shown in Fig. 2. In the seeds treated with artificial rainfall at 40 DAH, Shindongjin did not show a change in vigor or germination percentage regardless of rainfall treatment period. However, Hopum showed an approximately 10% decrease in vigor and a 10 to 20% decrease in germination compared to the non-treated seeds. When the artificial rainfall was applied at 50 DAH, the seed vigor and germination percentage decreased with increased rainfall duration in both cultivars. However, the decrease was much greater in Hopum than in SDJ.
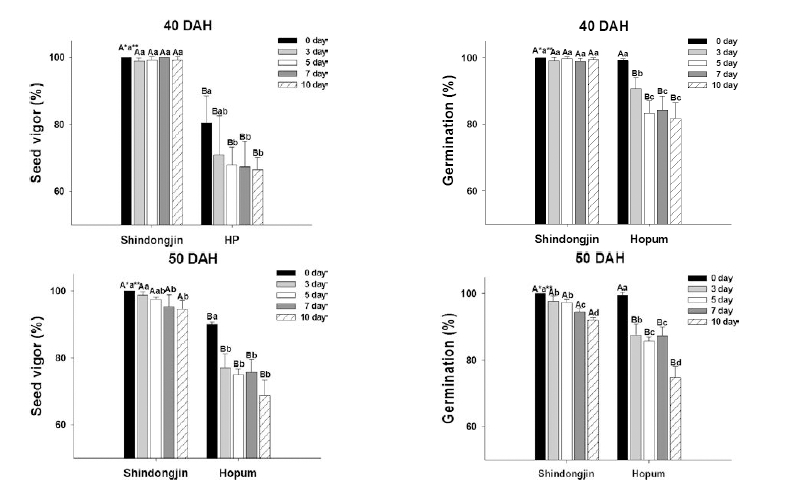
Fig. 2.
Seed vigor and germination (%) of two rice varieties after harvest. (*Different capital letters on the same date indicate a significant difference in means at α = 0.05 by t-test. **Different small letters for the same variety indicate a significant difference in means at α = 0.05 by Duncan’s Multiple Range Test (DMRT)).Seed longevity according to artificial rainfall period
The accelerated aging test was conducted with seeds treated with artificial rainfall during the ripening stage. The seed viabilities with different accelerated aging treatments from 0 to 30 days are shown in Table 2.
Table 2.
Seed viability tested after accelerated aging in two rice varieties following the rainfall treatment.
| Days of accelerated aging† | DAH | varieties | Days of rainfall treatment | ||||
|---|---|---|---|---|---|---|---|
| 0 | 3 | 5 | 7 | 10 | |||
| ---------------------------------------------- % ---------------------------------------------- | |||||||
| 0 | 40 | Shindongjin | 100.0 A*a** | 99.0 Aa | 99.3 Aa | 98.5 Aa | 98.8 Aa |
| Hopum | 99.3 ABa | 90.8 BCb | 84.3 Cc | 83.3 Dc | 81.8 Dc | ||
| 50 | Shindongjin | 100.0 Aa | 98.7 Ab | 97.3 Ab | 94.3 Bc | 92.0 Bd | |
| Hopum | 99.5 ABa | 87.5 Cb | 82.8 CDc | 80.3 Dc | 74.8 Ed | ||
| 5 | 40 | Shindongjin | 96.8 ABCa | 93.3 Bb | 92.3 Bbc | 91.4 BCbc | 89.5 BCc |
| Hopum | 93.5 CDa | 73.0 Eb | 62.0 Gc | 63.5 Fc | 47.5 GHd | ||
| 50 | Shindongjin | 96.3 BCa | 90.4 BCb | 88.2 Bc | 87.9 Cc | 85.2 CDd | |
| Hopum | 91.3 Da | 59.8 Gb | 62.8 Gb | 57.8 Gb | 45.0 Hc | ||
| 10 | 40 | Shindongjin | 95.3 Ca | 91.3 BCb | 89.3 Bc | 88.7 Cc | 85.5 CDd |
| Hopum | 83.8 Fa | 61.5 FGb | 50.7 Hc | 49.3 Hc | 35.0 Id | ||
| 50 | Shindongjin | 94.7 Ca | 90.1 BCb | 88.7 Bc | 88.5 Cc | 84.7 CDd | |
| Hopum | 82.5 Fa | 64.7 Fb | 68.0 Fb | 54.3 Gc | 46.0 Hd | ||
| 15 | 40 | Shindongjin | 88.0 Ea | 77.7 Db | 77.7 Eb | 70.7 Ec | 56.0 Fd |
| Hopum | 59.3 Ha | 47.0 Hb | 32.0 Jc | 15.0 Ld | 12.3 Kd | ||
| 50 | Shindongjin | 85.0 EFa | 77.7 Db | 79.5 DEc | 68.0 Ed | 51.2 Ge | |
| Hopum | 62.7 Ga | 46.0 Hb | 40.7 Ibc | 38.7 Ibc | 34.0 Ic | ||
| 20 | 40 | Shindongjin | 36.0 Ia | 34.0 Ia | 31.3 Ja | 29.0 Jab | 23.3 Jb |
| Hopum | 12.7 La | 13.3 KLa | 10.5 Na | 10.0 Ma | 9.3 Ka | ||
| 50 | Shindongjin | 32.8 Ja | 31.8 IJab | 30.1 Jb | 27.8 Jc | 24.3 Jd | |
| Hopum | 31.3 Ja | 29.3 Jab | 24.7 Kbc | 21.3 Kc | 12.7 Kd | ||
| 25 | 40 | Shindongjin | 18.9 Ka | 15.4 Kb | 14.5 Lb | 10.9 Mc | 11.5 Kc |
| Hopum | 6.7 Ma | 6.5 Ma | 3.4 Ob | 3.0 Nb | 2.9 Lb | ||
| 50 | Shindongjin | 17.9Ka | 14.3 KLb | 13.5 LMb | 10.2 Mc | 9.8 Kc | |
| Hopum | 10.6La | 10.9 La | 9.8 NMa | 7.6 Mb | 2.1 Lc | ||
| 30 | 40 | Shindongjin | 0.8 Na | 0.0 Na | 0.0 Oa | 0.0 Na | 0.0 La |
| Hopum | 1.3 Na | 0.0 Na | 0.0 Oa | 0.0 Na | 0.0 La | ||
| 50 | Shindongjin | 1.2 Na | 0.0 Na | 0.0 Oa | 0.0 Na | 0.0 La | |
| Hopum | 2.0 Na | 0.0 Na | 0.0 Oa | 0.0 Na | 0.0 La | ||
There was no cultivar viability difference in control seeds (non-rainfall treatment and non-accelerated aging seeds). However, accelerated aging differentiated seed longevity between the two cultivars. Cultivar differences were not observed up to five days of accelerated aging in non-artificial rainfall-treated seeds. Starting from ten days of accelerated aging, cultivar viability differences became clear; seed germination of Shindongjin was 10% higher than that of Hopum. After 15 days of accelerated aging, the viability difference between the two cultivars was much larger; Shindongjin showed 86.5% germination, but Hopum showed only 61.0%. In the seeds subjected to more than 20 days of accelerated aging, seed viability rapidly decreased in both cultivars, but the viability of Shindongjin was still higher than that of Hopum until 25 days of accelerated aging. At 30 days of accelerated aging, most seeds in both cultivars were non-viable.
Period of artificial rainfall affected seed longevity during accelerated aging. The viability of accelerated aging seeds decreased with prolonged rainfall period during the ripening stage. In the seeds subjected to only five days of accelerated aging, ten days of rainfall at the ripening stage decreased seed viability (average of 40 DAH and 50 DAH) from 96.6% to 87.4% in Shindongjin and from 92.4% to 46.3% in Hopum, respectively. In the case of 15 day- accelerated aging seeds, a ten day rainfall reduced seed viability from 86.5% to 53.6% in Shindongjin and from 61.0% to 23.2% in Hopum.
Concerning cultivar variation of seed longevity between Shindongjin and Hopum, the decrease in viability in response to rainfall treatment was observed to occur faster in Hopum than in Shindongjin. Shindongjin maintained its germination (>50%) ability for up to 15 days of accelerated aging regardless of DRT or DAH. However, Hopum dropped below 50% germination after as little as five days of accelerated aging in ten DRT-seeds and in 15 days of accelerated aging in three, five, and seven DRT-seeds.
Discussion
The two varieties used in this study showed regarding the artificial rainfall treatment time, Hopum showed similar pre-harvest sprouting responses to rainfall at 40 DAH and 50 DAH. In contrast, Shindongjin showed pre-harvest sprouting only at 50 DAH, and its germination percentage was much lower than that of Hopum. These results imply that Hopum is susceptible to unpredictable rainfall occurring after 40 DAH, and even a short duration of rain (three days) can cause pre-harvest sprouting.
Seed longevity was affected by rainfall during the ripening stage. Based on the accelerated aging test of rainfall-treated seeds, both cultivars demonstrated decreased longevity, and longer rainfall duration further reduced seed longevity. Lee et al. (2006) also reported that seed longevity and pre-harvest sprouting were linearly correlated, with a highly negative correlation coefficient. The reduced seed longevity in response to rainfall period was also faster in Hopum than Shindongjin.
In the seeds demonstrating pre-harvest sprouting, the germination process was disrupted by desiccation after radicle emergence. In this case, grains maintain their viability, but seed longevity is reduced dramatically. If damp conditions in the field persist longer, the germination process may proceed toward a point of no return, beyond which the embryo loses desiccation tolerance (Rodríguez et al., 2001; Gualano and Benech-Arnold, 2009; Gerjets et al., 2010). Even mild levels of sprouting damage can substantially reduce grain storage life (Li et al., 2004).
Recently, there were more rainy days in Korea during the rice ripening stage. Consequently, many rice cultivars failed to achieve good seedling establishment because of poor germination and low viability of rice seed. Among them, Hopum was one of the varieties that failed to germinate and grow in the field (Kim et al., 2008; Korean Rural Economic Institute, 2012; RDA, 2013). Therefore, the present study verified that the varietal difference between Hopum and Shindongjin, as related to pre-harvest sprouting response to rainfall during the ripening stage, could make difference in seed viability and longevity after harvest and storage. Seed dormancy and pre-harvest sprouting are expressed as quantitative traits that are strongly influenced by environment, as well as interactions between genotype and environment (Liu et al., 2008). Early disruption of seed dormancy in wet or humid conditions prior to harvest generally causes pre-harvest sprouting, although cultivars with strong seed dormancy are more resistant to pre-harvest sprouting. Our results show that rice seeds of pre-harvest sprouting rapidly lose viability during storage. pre-harvest sprouting tolerance is related to seed viability and longevity during storage. The long-term solution to pre-harvest sprouting relies on the development of cultivars that are able to tolerate pre-harvest sprouting during the maturation period and to resist degeneration after harvest and climate change (Li et al., 2004; Gao et al., 2008). In conclusion, rainfall during the ripening stage induced not only pre-harvest sprouting, but also reduced seed quality and longevity during storage, which varied between two cultivars.
