

Grains, the seeds of many grasses, are considered a main source of carbohydrates, minerals and vitamins in a balanced diet. They also provide fiber, which has various health promoting functions such as anti-diabetic effect, cholesterol lowering effect, and prevention of heart disease (Brown et al., 1999; Marlett et al., 2002; Pereira et al., 2004).
Furthermore, seeds play a crucial role in the life cycle of plants; because they are essential for plant reproduction (Yang et al., 2009). Cereal crops are cultivated mainly for the stored protein, oil, and carbohydrates that eventually accumulated in seeds. Seeds storage protein (SSP) is the main source of carbon, nitrogen, sulfur triglycerides and carbohydrate reserves (Herman and Larkins 1999). SSPs are convinced to be an authentic source as plant proteins that consumed by humans. During the posterior stages of seed development, SSPs are accumulated in seed development and degraded during germination (Jha et al., 2012). Moreover, SSPs play an important role for the development of seedling growth and also important source of protein for humans and animals (Foley et al., 2011).
The proso millet (Panicum miliaceum L.) is a cereal plant cultivated for its grain, mostly in Asia and North America. It is a warm-season grass with a short growing season and low moisture requirement that is capable of producing food or feed where other grain crops would fail (Baltensperger et al., 1995; Helm et al., 1990). Millet is a general term for a wide range of small seeded cereals (Marçon 1994). The millets of the Poaceae (Gramineae) are small seeded annual grasses, including a wide range of cereals, that may be used both as grain and for forage. Proso millet (2n=4x=36), an allotetraploid self-pollinated cereal with highest water-use efficiency, is the best rotational crop in dryland production system in the semi-arid high plains of the USA and elsewhere in the world (Rajput 2015).
Previous report revealed that millet proteins are good sources of essential amino acids except lysine and threonine, but are relatively high in methionine (Mal et al., 2010). In proso millet grains, protein content is comparable with that of wheat, but the share of essential amino acids (leucine, isoleucine and methionine) is substantially higher in proso millet (Saleh et al., 2013). Proteomic approaches has gained significant attention as a tool of protein identification in plants and many proteomic studies have been conducted on some grains such as soybean (Koo et al., 2011), pea (Bourgeois et al., 2009), peanut (Kottapalli et al., 2008), salicornia (Jha et al., 2012), subclover (Sahnoun-Abid et al., 2009), lupin (Islam et al., 2012), rapeseed (Hajduch et al., 2006), medicago (Gallardo et al., 2003), arabidopsis (Gallardo et al., 2001), wheat (Islam et al., 2002; Majoul et al., 2003; Skylas et al., 2001), sorghum (Mesa-Stonestreet et al., 2010) and barley (Finnie et al., 2004). However, proteome profiling have conducted in the germinating seeds of soybean through one-dimensional gel electrophoresis followed by liquid chromatography combined with tandem mass spectrometry and confirmed 764 proteins belonging to 14 functional groups as well as metabolism related proteins in the largest group (Han et al., 2013). In pea, 156 identified proteins have provided a fine dissection of the seed storage protein which ultimately exposing a large diversity of storage proteins (Bourgeois et al., 2009).
Although, few preliminary reports of millet using 2-D gel electrophoresis have been employed by time, especially towards salinity stress (Veeranagamallaiah et al., 2008), there is still lack of study on protein profiling using electrophoresis with MALDI TOF/TOF MS. In the present study, a high throughput proteome technique was employed for establishing a comprehensive seed proteomes profile from proso millet (Panicum miliaceum L.). The proteomic profiling of Proso millet seeds were analyzed using 2D IEF/PAGE electrophoresis with MALDI-TOF/TOF MS for obtaining a reference map, understanding the functional characteristics of the identified proteins and identifying cultivar-specific protein expression changes.
MATERIALS AND METHODS
Experimental materials
Four different cultivars (Leebaekchal, Manhongchal, Miryang 7 and Miyang 8) of proso millet (Panicum miliaceum L.), harvested in 2013, were obtained from Rural Development Administration (RDA), Miryang, Korea, and stored in a refrigerator at 4°C prior to analysis.
Extraction of proteins
Seed proteins were extracted according to previously described methods (Kamal et al., 2011) with a slight modification. Proso millet seeds (500 mg) were ground in a mortar with liquid nitrogen. The samples (seeds) were mixed with the solution I (10% TCA in acetone containing 0.07% 2-mercaptoethanol) and then sonicated for 10 min. After sonication, the suspensions were centrifuged at 20000 x g for 5 min at 4°C. The precipitates (residues) were washed twice with the solution II (100% acetone containing 0.07% 2-mercaptoethanol), and then vacuum-dried to dryness at 25°C by speed vac concentrator. The dried sample was suspended in lysis buffer (7 M urea, 2 M thiourea, 5% CHAPS, and 2 mM tributylphosphine). After incubation, the supernatants were recovered by centrifuging (20,000 x g, for 20 min at 4°C) and stored at -80°C for the analyses. The protein concentration in samples was determined by Bradford assay (Bradford 1976).
Gel electrophoresis and image analysis
Two-dimensional electrophoresis of proteins in the proso millet was performed according to the previously described protocol (O'Farrell 1975) with minor modification. Sample solutions (400 μg) were loaded onto the acidic side of the IEF tube gels (11 cm × 2 mm), and an IEF gel was adopted specifically for pH range 3 to 10 (carrier ampholyte) in addition to the acidic and basic ranges. SDS-PAGE in the second dimension (Nihon Eido, Tokyo, Japan) was executed with 12% separation and 5% stacking gels with 13 cm x 13 cm gel plates. Protein spots in the 2-DE gels were visualized by Coomassie Brilliant Blue (G-250) staining. Each biological sample was carried out three times and the visualized gels were selected for image analysis in each replication. 2-DE gels were scanned with an image scanner (HP Scanjet G4010, CA, USA; 300 dpi, 32 bits per pixel). Computer assisted 2-DE image analysis was performed with Progenesis SameSpot software version 3.0 (Nonlinear Dynamics Ltd, Durham, NC, USA).
In-gel digestion
CBB-stained gel slices were washed several times with 30% methanol until the colors were completely removed. Then the gel slices were destained with 10 mM (NH4) HCO3 in 50% ACN (Acetonitrile), squeezed for 10 min with 100% ACN and dried by vacuum centrifugation. After destaining steps, the gel slices were reduced with 10 mM DTT in 100 mM (NH4)HCO3 at 56°C for 1hr and then alkylated with 55 mM iodoacetamide (IAA) in 100 mM (NH4)HCO3 in the dark for 40min. Then the gel slices were digested with 50 μL trypsin buffer (Promega Corporation, Madison, WI 53711- 5399, USA) and incubated at 37°C for 16 hours. After digestion steps, the peptides were extracted with 50 mM ABC (Ammonium Bi-Carbonate) and repeated these steps several times with a solution containing 0.1% formic acid in 50% ACN (Acetonitrile) until 200~250 μL. The solution containing eluted peptides was concentrated up to drying by vacuum centrifugation and the resultant extracts were confirmed by MALDI-TOF-TOF mass spectrometry.
Mass spectrometry (MALDI-TOF-TOF/MS) analysis
Selected spots from 2-DE gels were analyzed to evaluate the compatibility between protein extraction and mass spectrometry as well as to disclose the protein classes that populate each 2-DE gels. Mass spectra were acquired in an ABI 4700 Proteomics Analyzer (Applied Biosystems) using 3, 5-dimethoxy- 4-hydroxycinnamic acid as matrix and the resulting data by the GPS Explorer package (Applied Biosystems). Peptides were dissolved in 0.5% (v/v) trifluoroacetic acid (TFA) and desalted with a ZipTip C18 (Millipore, Bedford, MA, USA). Those purified peptides were then eluted directly onto a MALDI plate by using an α-cyano-4- hydroxycinnamic acid (CHCA) matrix solution [10 mg per cm3 of CHCA in 0.5% (v/v) TFA + 50% (v/v) acetonitrile; 1:1]. All mass spectra were acquired in the reflection mode with 0 - 4000 m/z by a 4700 proteomics analyzer (Applied Bio-systems, Framingham, MA, USA). External calibration was performed using a standard peptide mixture of des-Arg bradykinin, angiotensin, Glufibrinopeptide B, adrenocorticotropic hormone (ACTH) clip 1-17, ACTH clip 18-39, and ACTH clip 7-38.
Bioinformatics analysis of proteins
To identify the peptides, acquired MS/MS spectra were evaluated using Mascot Generic File (MGF) with an in-house licensed MASCOT search engine (Mascot v. 2.3.01, Matrix Science, London, UK) against the viridiplantae within the NCBInr database. MASCOT was used with the monoisotopic mass selected, a peptide mass tolerance of 50 ppm, and a fragment iron mass tolerance of 2 Da. The instrument setting was specified as MALDI-TOF/TOF. The carbamidomethylation of cysteines was set as a fixed modification whereas the oxidation of methionines was set as a variable modification. Trypsin was specified as the proteolytic enzyme with one potential missed cleavage. All proteins identified by high-scoring peptides were considered true matches, and at least two peptide matches. Protein hits were validated if the identification involved at least 10 top-ranking peptides with p < 0.05 and peptide scores > 34, and also selected false positive rate < 0.05. When those peptides matched multiple members of a protein family, the presented protein was selected based on the highest score and the greatest number of matching peptides.
Proteins identified from the present study were grouped into functional categories based on gene ontology using data available on the protein information resources (PIR) (http://pir.georgetown.edu) as well as literature sources.
RESULTS AND DISCUSSION
Separation of millet seeds by 2-DE
Protein samples were extracted from the matural proso millet seeds of the four cultivars (Leebaekchal, Manhongchal, Miryang 7 and Miyang 8) and were analyzed to investigate the seed storage proteins using proteomic techniques. The 1152 reproducible protein spots were detected in 2D gel, which were mostly placed in the center of 2D gels (Fig. 1). However, the pattern of 26 protein spots was significantly different between the four cultivars when the difference threshold was set to more than 1.5 fold (Fig. 3). Among the 26 identified proteins, 2 proteins were up-regulated against all millet cultivars. Thirteen proteins (13) were up-related while 11 proteins were down-regulated against two cultivars (Table 1). Interestingly, some of the differentiating proteins identified as a chain form in the gels with similar molecular weights but different pI values. The pI of 13 protein spots was between 5 and 6 accounted for 50% of the total number of protein spots (Fig. 2A). In addition, the molecular mass of ~35% of the proteins was between 20 kDa and 30 kDa (Fig. 2B).

Fig. 1
Representative gel images from the 2-D electrophoresis analysis of Leebaekchal (A), Manhongchal (B), Miryang 7 (C), and Miyang 8 (D) proso millet seeds. The seeds were fractionated using the TCA-acetone precipitation method, as described in the Materials and Methods section. Proteins were extracted from the seeds of the four proso millet cultivars. For iso-electric focusing, 400 μg samples were loaded onto the acidic side of IEF tube gels (11 cm × 2 mm), which were pre-run at 150 V for 1 h, 300 V for 1 h, and 500 V for 16 h for the first dimension. Second-dimensional electrophoresis (Nihon Eido, Tokyo, Japan) was performed with 12% separation and 5% stacking gels with 13 cm × 13-cm gel plates and then stained with CBB. The differentially expressed protein spots (>1.5-fold difference) are indicated using circles on the 2-D gel map and are statistically significant to a level of 95% per group (Student’s t-test) using biological and analytical replicates (n = 3). The MW of each protein was determined using standard markers.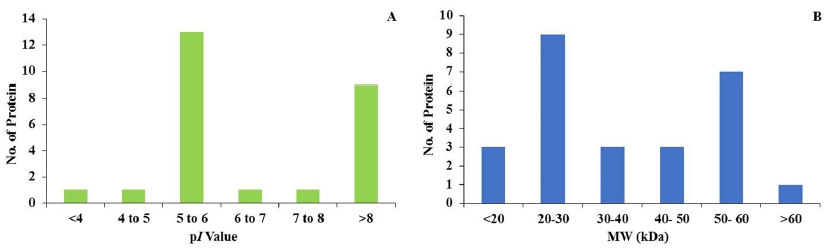
Fig. 2
Differential (A) iso-electric point (pI) and (B) molecular weight (kDa) of the identified proteins from Leebaekchal, Manhongchal, Miryang 7, and Miyang 8 cultivars of proso millet (Panicum miliaceum).
Fig. 3
Relative protein intensities of differentially expressed proteins from different proso millet cultivars. The proteins were extracted from seeds of the four proso millet cultivars. Results are presented as mean ± SE values of the relative protein intensities of 3 biological replicates. The gels were stained with CBB, and differential abundance of the proteins was determined using Progenesis SameSpot software.Table 1
Differentially changed proteins identified from seeds of Leebaekchal, Manhongchal, Miryang 7, and Miyang 8 cultivars of Panicum miliaceum by using MALDI-TOF-TOF mass spectrometry.
| SNa | PAb | Protein Description | PSc | MWd | pIe | PMf | PCg | Cultivar-Specific Differential Protein Expression | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leebaekchal | Manhongchal | Miryang7 | Miryang8 | ||||||||
| Glycolysis | |||||||||||
| 122 | gi|497503398 | L-carnitine dehydrogenase | 50 | 52114 | 7.6 | 8 | 26 | Control | X | ✓ | X |
| 294 | gi|90110845 | Enolase | 105 | 48285 | 5.6 | 10 | 39 | Control | ✓ | X | ✓ |
| Stress response | |||||||||||
| 160 | gi|527187624 | Heat shock protein 70 | 98 | 71617 | 5.1 | 15 | 26 | Control | ✓ | X | X |
| 255 | Q2QU06 | 60 kDa chaperonin alpha subunit | 188 | 61150 | 5.1 | 10 | 42 | Control | X | X | ✓ |
| 256 | Q7X9A7 | 60 kDa chaperonin alpha subunit | 175 | 61477 | 5.2 | 11 | 52 | Control | ✓ | ✓ | ✓ |
| 404 | gi|48716124 | Heat shock protein | 163 | 43113 | 5.1 | 14 | 19 | Control | X | X | ✓ |
| Metabolism | |||||||||||
| 179 | gi|512440358 | GMP synthase | 61 | 59106 | 5.7 | 13 | 18 | Control | ✓ | X | X |
| 322 | gi|546164593 | Uncharacterized protein | 67 | 41967 | 9.2 | 17 | 29 | Control | ✓ | X | ✓ |
| 431 | gi|447018072 | Uncharacterized protein | 64 | 33647 | 9.3 | 10 | 74 | Control | X | ✓ | ✓ |
| 529 | gi|315426860 | methyltransferase | 51 | 19727 | 10.1 | 9 | 29 | Control | ✓ | ✓ | ✓ |
| 952 | A4S6Y3 | Uncharacterized protein | 46 | 21594 | 8.8 | 9 | 40 | Control | X | ✓ | X |
| Starch metabolism | |||||||||||
| 325 | A7XAQ5 | Glucose-1-phosphate adenylyltransferase | 58 | 57690 | 4.6 | 16 | 14 | Control | ✓ | X | ✓ |
| Polysaccharide metabolism | |||||||||||
| 400 | gi|324021404 | Starch synthase, chloroplastic/amyloplastic | 285 | 54585 | 6 | 20 | 32 | Control | ✓ | X | ✓ |
| 453 | gi|324021404 | Starch synthase, chloroplastic/amyloplastic | 230 | 54585 | 6 | 17 | 32 | Control | X | ✓ | ✓ |
| Transcription | |||||||||||
| 525 | gi|495043419 | XRE family transcriptional regulator | 60 | 24849 | 6.3 | 9 | 23 | Control | X | ✓ | ✓ |
| Biosynthetic pathway | |||||||||||
| 628 | gi|24374432 | cysteine synthase CysK | 55 | 34456 | 5.3 | 6 | 20 | Control | ✓ | ✓ | X |
| 653 | gi|496152858 | ATP phosphoribosyltransferase | 54 | 25027 | 10.8 | 8 | 35 | Control | ✓ | X | X |
| Mitotic nuclear division | |||||||||||
| 627 | gi|540385101 | Kinetochore protein | 59 | 36866 | 5.2 | 6 | 19 | Control | X | ✓ | X |
| Protein Repair | |||||||||||
| 669 | gi|490733491 | Methionine-R-sulfoxide reductase | 40 | 15782 | 3.8 | 3 | 97 | Control | ✓ | ✓ | X |
| Transduction | |||||||||||
| 699 | gi|406865556 | Rho3 protein | 48 | 23420 | 9.7 | 4 | 13 | Control | X | ✓ | X |
| Pathogenesis | |||||||||||
| 951 | gi|323393606 | erythrocyte membrane protein 1 | 50 | 20974 | 11.6 | 9 | 31 | Control | X | ✓ | ✓ |
| Unknown functions | |||||||||||
| 664 | gi|493302409 | Uncharacterized protein | 42 | 34510 | 8.5 | 6 | 32 | Control | ✓ | X | ✓ |
| 688 | gi|23490617 | Uncharacterized protein | 33 | 2917 | 5.8 | 2 | 97 | Control | ✓ | ✓ | X |
| 706 | gi|29345740 | ketoisovalerate oxidoreductase subunit | 40 | 26134 | 5.4 | 6 | 20 | Control | X | ✓ | X |
| 879 | gi|170584994 | Zinc finger | 40 | 19300 | 5.3 | 5 | 74 | Control | ✓ | X | X |
| 984 | B9SSH4 | Calcium ion binding protein, putative | 42 | 15600 | 12 | 5 | 55 | Control | X | ✓ | ✓ |
a Represents the spot number on the master gel
b
Accession number according to the UniProt database
c
Score of the identified peptides
d
Molecular weight of the identified proteins
e
Iso-electric point of the identified proteins
f
Number of matched peptides, the proteins with >2 matched peptides were considered
g
Protein coverage, the proteins with less than 10% sequence coverage was excluded from the result
The symbol “X” denotes the absence of protein expression and “⚐” denotes the presence of protein expressio
Functional classification of identified proteins
The 26 differentially expressed protein spots in the proso millet seeds were identified by MASCOT DB and classified into the functional categories. Gene ontology categories were assigned to all 26 proteins according to their molecular function, cellular component and biological processes (Fig. 4 A-C).

Fig. 4
Protein encoding gene function of 26 differentially expressed proteins from proso millet seeds. Frequency distribution of the identified proteins within functional categories was based on molecular functions (A), cellular components (B), and biological processes (C). Classifications were made using iProClass databases, and assignment by function was based on gene ontology. Blue and red bars indicate induced and reduced proteins, respectively.The proteins were classified into 8 groups according to molecular function. The nucleic binding group has the greatest number of the identified proteins followed by transferase activity and ion binding (Fig. 4A). The identified proteins were classified into 5 categories according to cellular location (localization). Most of the proteins were localized in the membrane, plastids and cytoplasm (Fig. 4B).
Based on biological function, the identified proteins were classified into the following 10 biological process categories: glycolysis [L-carnitine dehydrogenase (spot 122), enolase (spot 294),]; stress response [Heat shock protein (spot 160), 60 kDa chaperonin alpha subunit (spot 255), 60 kDa chaperonin alpha subunit (spot 256), heat shock protein (spot 404)]; metabolism [GMP synthase (spot 179), uncharacterized protein (spot 322), uncharacterized protein (spot 431), glucose-1-phosphate adenylyltransferase (spot 325), starch synthase, chloroplastic/amyloplastic (spot 400), starch synthase, chloroplastic/amyloplastic (spot 453), methyltransferase (spot 529), uncharacterized protein (spot 952)]; mitotic nuclear division [kinetochore protein (spot 627)]; transcription [XRE family transcriptional regulator (spot 525)]; Biosynthetic pathway [ATP phosphoribosyltransferase (spot 653), Cysteine synthase CysK (spot 653)]; protein repair [methionine-R-sulfoxide reductase (spot 669)]; transduction [Rho3 protein (spot 699)]; pathogenesis [6-phosphogluconate dehydrogenase protein (spot 951)]; and unknown functions [uncharacterized protein (spot 664), uncharacterized protein (spot 688), ketoisovalerate oxidoreductase (spot 706), zinc finger (spot 879), calcium ion binding protein, putative (spot 984)]. Among the 10 biological process categories, methbolism group has the largest number of proteins followed by stress response (Fig. 4C).
Changes in protein expression patterns in millet seeds
Proteomic technologies are the most widely applied for protein identification in cereals (Chi et al., 2010; Chitteti and Peng 2007; Ding et al., 2012; Fan et al., 2011; He et al., 2011; Nwugo and Huerta 2010; Torabi et al., 2009; Yang et al., 2007a; Yang et al., 2006). In this study, 2-DE combined with MALDI-TOF-MS was carried out for protein profiling of proso millet seeds. The identified proteins were classified based on the biological functions; and their expressions were ellustrated in the Fig. 5. Several unknown proteins were not identified due to the lack of genomic sequencing. Some of the identified millet seed protein species associated with primary biological processes are discussed below.
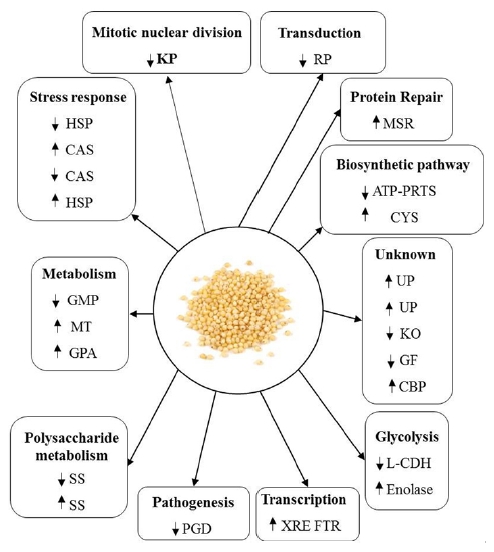
Fig. 5
An overview of the differentially expressed proteins of Panicum miliaceum seeds. L-CDH: L-carnitine dehydrogenase; HSP: Heat-shock protein 70; GMP: GMP synthase; CAS: 60-kDa chaperonin alpha subunit; UP: Uncharacterized protein; GPA: Glucose-1-phosphate adenylyltransferase; SS: Starch synthase, chloroplastic/ amyloplastic; XRE FTR: XRE family transcriptional regulator; MT: Methyltransferase; KP: Kinetochore protein; Cys: Cysteine synthase CysK; ATP-PRTase: ATP phosphoribosyltransferase; MSR: Methionine-Rsulfoxide reductase; RP: Rho3 protein; KO: Ketoisovalerate oxidoreductase; ZF: Zinc finger; PGD: 6-Phosphogluconate dehydrogenase protein; CBP: Calcium ion-binding protein, putative.Proteins involved in metabolism
Metabolism-related proteins play an important role to maintain seed vigor and are mainly associated with carbohydrate metabolic pathways including glycolysis and the TCA cycle (He et al., 2011). A large number of metabolism-related proteins were found in proso millet seeds. The expression of these proteins was significantly changed by storage.
The abundance of L-carnitine dehydrogenase (Table 1, spot 122) was significantly increased in the Miryang7 cultivar, but decreased in the Manhongchal and Miryang8 cultivars. However, enolase (Table 1, spot 294), which was remarkably identified in the proso millet seeds, was down-regulated in the Miryang7 cultivar but up-regulated in the Manhongchal and Miryang8 during seed storage. Many plastidial glycolytic and Calvin cycle were activated during seed filling in the four oil seeds in Brassica napus L., except PGAM and enolase. These activities are required for the carbon dioxide recycling in plant (Schwender et al., 2004). Some studies reported that enolases were decreased during the development and germination in plant seeds (He and Yang 2013; Li et al., 2012).
Guanosine 5’-phosphate synthase (Table 1, spot 179) contributing the formation of GMP and PPi from xantosine-5’P was down-regulated in the Miryang7 and Miryang8 cultivars, but up-regulated in the Manhongchal cultivar during the storage. It appears that GMP synthase substrate (xanthosine- 5’P) could derive from biochemical routes involving a pool of xantine and xantosine-triP or-tetraPi in a salvage pathway (Gallo et al., 2010).
The upregulation of methyltransferase (Table 1, spot 529) was found towards all cultivars of proso millet seeds. Oge et al. (2008) exhibited that methyltransferase induces longevity and germination vigour in Arabidopsis.
Proteins involved in starch metabolism
Three starch metabolism-related proteins such as glucose-1- phosphate adenylyltransferase (Table 1, spot 325), starch synthase, chloroplastic/amyloplastic (Table 1, spots 400, 453) are found in the proso millet seeds. The level of glucose-1- phosphate adenylyltransferase was increased in the Manhongchal and Miryang8 cultivars but decreased in the Miryang 7 cultivar. Starch synthase, chloroplastic/amyloplastic was up-regulated in the Manhongchal and Miryang8 cultivars but down-regulated in the Miryang7. Emes et al. (2003) demonstrated that grain yield is largely affected by starch accumulation during the grain filling phase in cereals. Starch biosynthesis is initiated with ADP-glucose produced by glucose- 1-phosphate adenylyltransferase (ADP-glucose pyrophosphorylase/ AGPase) reaction. AGPase, located in plastids, is a key regulatory enzyme in starch biosynthesis (Smidansky et al., 2007). Granule-bound starch synthase (GBSS), one of the key enzymes in the starch biosynthesis, converts UDP-glucose and ADP-glucose in plant tissue (Macdonald and Preiss 1983).
Proteins involved in stress response
Several proteins including heat shock protein 70 (Table 1, spots 160, 404), 60 kDa chaperonin alpha subunit (Table 1, spots 255, 256) were found in the proso millet seeds. Heat shock factor binding protein (HSP, spot 160) was up-regulated in the Manhongchal cultivar but down-regulated in the Miryang7 and Miryang8 cultivars during storage of proso millet seeds. HSPs a kind of molecular chaperone, are involved in the reparing of stress-damaged proteins through the protein re-naturation (Sun et al., 2002; Vierling 1991). The expression of heat-shock proteins (HSPs) are associated with desiccation. Guo et al. (2007) suggested that HSP 70 plays an important role in cellular redox regulation . Some studies have demonstrated that several genes, which involved with HSPs, expressed during seed development under heat stress (Crosatti et al., 1999; Wehmeyer and Vierling 2000). Abundance of two HSPs proteins, HSP70 (Table 1, spot 160) and heat shock cognate 60 kDa chaperonin alpha subunit (Table 1, spot 255) were the major HSP proteins in the proso millet seeds that were significantly increased during storage (Table 1, Fig. 3). HSPs is thought to function by restoring protein structure and prohibiting protein denaturation (Wang et al., 2004). Although the expression of HSP 70 decreased during germinatiom, it was still remained in Arabidopsis (Gallardo et al., 2001). Georgopoulos and Welch (1993) reported similar results showing that the expression of these HSP proteins were increased in response to heat stress. Some researchers demonstrated that the desiccation, water stress and heat shock reduced the abundance of these proteins in the creeping bent grass cultivars, which are contrary to our results (Xu and Huang, 2010; Sano et al., 2013). Yacoubi et al.,et al. (2011) reported that the HSP 70 and GroEL-like chaperone were up-regulated in Medicago sativa during germination.
Proteins involved in transcription
XRE family transcriptional regulator (Table 1, spot 525) was up-regulated in the Miryang7 and Miryang8 cultivars but down-regulated in the Manhongchal cultivar of the proso millet seeds. However, Rajjou et al. (2004) reported that XRE family transcriptional regulator was up-regulated in Arabidopsis during storage.
Proteins involved in cysteine biosynthetic pathway
Cysteine synthase, which required for cysteine biosynthesis in plants is one of the most important enzymes in sulfur assimilation to lead the production of antioxidants and metal chelators, such as glu-tathione, metallothionein, and phytochelatin (Yang et al., 2007b). Cysteine synthase CysK (Table 1, spot 628) was found to be increased towards Manhongchal and Miryang7 cultivars but decreased against Miryang8 cultivar in the proso millet seeds. Cysteine is first reduced by a sulphur-containing compound and carried out as a sulphur donor for Met (Chronis and Krishnan 2003). Ado-Met synthase is a key enzyme in plant metabolism, catalysing the biosynthesis from Met to Ado-Met. Ado-Met is not only the primary methylgroup donor but also a source of amino-alkyl and amino groups for the biosynthesis of amino acids, polyamines, and hormone ethylene. It also could provide the methylene group used in the biosynthesis of fatty acids that accumulate in many types of seeds (Bouvier et al., 2006).
Proteins with miscellaneous functions
In the present study, several proteins such as kinetochore protein (Table 1, spot 627), ATP phosphoribosyltransferase (Table 1, spot 653), methionine-R-sulfoxide reductase (Table 1, spot 669), Rho3 protein (Table 1, spot 699), 6-phosphogluconate dehydrogenase protein (Table 1, spot 951) were found in the proso millet seeds and were differentially expressed during seed storage.
Several unknown proteins in MASCOT DB were up-regulated (Table 1, spots 664, 688, 984) or down-regulated (Table 1, spots 706, 879) during seed storage of the proso millet seeds. Although this study successfully demonstrates the protein profile in the proso millet seeds, there is still lack of study on the function and origin of some proteins. Further studies are needed to understand their origin and function.
CONCLUSION
The protein profiling of proso millet seeds was performed using high throughput (2-D) IEF-PAGE with MALDI-TOF/TOF mass spectrometer. This study was an initial proteomic investigation of the four Korean cultivars of proso millet seeds and clearly domonstrates the proteome profiling of the proso millet. The 26 protein spots (> 1.5 folds) were found (visualized) in 2-D gel (stained with CBB), and then each protein (spot) was analyzed (identified) by mass spectrometer. The results obtained from the present study suggests that the changes of the differentially expressed proteins from the various millet cultivars are cultivar-specific. Most of them were involved in the starch metabolism, stress response and biosynthetic pathway. However several protein spot were not identified by MASCOT search engine. Further studies will focus on the identification of unknown proteins and changing mechanism during storage in the proso millet seeds.
