

콩(Glycine max L.)은 동아시아가 원산지로서 우리나라에 서는 중요한 식량작물의 하나로 재배되어 왔으며 필수 3대 영양소 중에서 가장 중요한 단백질이 40%로 우수한 단백질 공급원으로써 이용되어 왔다(Rahman et al., 2011). 콩의 주 요 종실 저장단백질은 총 단백질의 약 70~80%인 salt-soluble globulins로(Nielsen, 1996; Krishnan, 2000), 초기생육과정 에서 필요한 아미노산을 공급하기 때문에 종자가 발아하는 동안에 소모된다(Krishnan, 2002). 자엽에서 총 단백질의 축 적은 전반적으로 개화 후 15일부터 성숙기까지 이루어지며 특히 개화 후 25~40일 사이에 단백질의 축적비율이 매우 급 격하게 증가된다. β-conglycinin과 glycinin은 자엽에 존재 하는 저장단백질로 이 두 가지가 가장 풍부한 그룹이다 (Krishnan, 2002). 7S 단백질인 β-conglycinin은 종자가 발 달하는 동안에 α'-subunit, α-subunit 그리고 β-subunit으로 구성된 삼량체이다. β-subunit이 개화 후 5~7일 사이에 축 적되는 동안 α'-, α-subunits은 개화 후 18~20일 사이에 자엽 에 축적된다(Gayler and Sykes, 1981). 콩 종실의 발달 과정 을 보면 종실이 약 2 mm 일 때인 형태학상의 기간은 개화 후 2주이며 세포분열시기는 개화 후 3, 4주 그리고 세포 확 장 시기는 개화 후 5, 6주이다. 그러나 이 시기는 초기 배발 생 또는 종실이 성숙한 시기는 아니다(Mienke et al., 1981). 콩 종실이 발달하는 동안에 저장단백질은 개화 후 2, 3주에 서 유사한 발현양상을 보였으며 4, 5, 6주에서는 종실 저장 단백질의 함량이 약 35, 53, 60%로 증가되는 경향을 보였다 (Hajduch et al., 2005).
같은 식물 종이 동일 개체라 하더라도 생육단계화 조직기 관에 따라서 발현하는 단백질의 종류는 다르다. 이것은 게 놈 DNA의 염기서열이 동일종 내에서는 같지만 생육조건에 따라서 단백질의 발현은 다양한 변이를 나타내기 때문이다 (Woo et al., 2003). 따라서 본 연구에서는 등숙(week after flowering; waf)이 진행됨에 따라 콩 종실 저장단백질 발현 양상을 비교하여 등숙간 콩 종실 저장단백질 발현양상의 차 이를 구명하고 질량분석을 통하여 단백질의 기능을 구명하 였다.
재료 및 방법
식물재료
개화 후 등숙 과정에 따른 콩 종실 저장단백질의 발현양 상의 차이를 구명하기 위하여 태광콩을 식물재료로 사용하 였다. 태광콩을 충북대학교 부속농장 전작포장에 파종하였 으며. 시비는 콩 복비 50 k/10a (N : P2O5 : K2O = 3 : 3 : 3.4 kg/10a)을 전량기비로 하였으며, 기타 재배관리는 콩 표준재배법에 준하였다. 개화 후 1주 간격으로 1주에서 6주 까지 종실을 채취하여 -20°C에서 보관하였다가 본 실험에 사용하였다.
개화 후 시기별 단백질 추출 및 함량분석
이차원전기영동을 위한 단백질 추출을 위하여 개화 후 시기 별로 채집된 시료의 지방을 제거하기 위하여 85%의 n-hexane 을 사용하였다. 지방을 제거한 시료 0.1 g에 lysis buffer [4.8 g Urea, 0.2 ml NP-40, 0.2 ml Ampholyte pH 3.5-10, 0.5 ml 2-mercaptethanol, 0.5 g PVP-40/10 ml] 1 ml를 첨가하여 막대사발에서 혼합한 후 15,000 rpm에서 15분간 원심분리 하여 상등액을 취하고, 상등액을 분석하였다.
단백질 함량 분석은 Bradford (1976)의 방법에 595 nm에 서 분광광도계(SHIMADZU, UV-1700)를 사용하여 측정하 였다.
이차원전기영동 및 이미지 분석
이차원전기영동(O'Farrell, 1975)은 넓은 등전점영역의 단 백질을 높게 분리할 수 있는 능력을 가지고 있는 일차원(등 전점) 전기영동 NA-1313 (Nihon-Eido)과 이차원 전기영동 은 NA-1130 (Nihon-Eido)을 사용하였다. 일차원전기영동은 등전점전기영동으로 사용된 겔의 조성은 4.8 g Urea, 1.6 ml 30% Acrylamide solution, 2.84 ml DDW, 0.2 ml 10% NP-40, 0.25 ml Ampholyte pH 3.5-10, 0.25 ml Ampholyte pH 5-8, 15 μl 10% APS, 10 μl TEMED이다. 일차원 전기영동 후 평형화가 끝난 겔을 이차원전기영동을 하였다. 이차원전기 영동은 12% separating gel (9.6 ml DDW, 17.7 ml 30.0% acrylamide solution, 16.4 ml 1.0M Tris-HCl (pH 8.8), 300 μl 10% Ammonium persulfate, 50 μl TEMED)과 5% Stacking gel (2.0 ml DDW, 1.0 ml 30.0% acrylamide solution, 3.0 ml 0.25M Tris-HCl (pH 6.8), 30 μl 10% Ammonium persulfate, 20 μl TEMED)을 사용하였으며 수행하였다. 전기영동이 끝 난 후 겔은 은 염색법을 사용하였다. 은염색의 경우에는 Amersham사의 PlusOne Silver Staining Kit, Protein을 사 용하여 염색을 하였다. 이차원전기영동의 결과는 컴퓨터 프 로그램(Image analysis; nonlinear사의 Progenesis Worstation) 을 사용하여 겔상에서 단백질 spot의 변화를 비교분석하였다.
질량분석을 이용한 단백질 동정
이차원전기영동 후 겔에서 원하는 spot을 적당한 크기(1 mm X 1 mm)로 잘라서 e-tube에 넣었다. 겔을 탈색하기 위 해서 30mM Potassium ferricyanide/100mM sodium thiosulfate (1:1[V/V]) 100 μl를 첨가하여 흔든 후 원심분리를 실시한 후 상등액을 버렸다. 3차 증류수를 100 μl를 첨가한 후 원 심분리를 하여 다시 상등액을 버렸다. 이 과정을 3번 반복 하였다. 100% ACN 100 μl를 첨가하여 5분 동안 섞은 후 원심분리하여 상등액을 버렸다. 원심분리건조기에서 10분정 도 건조시켰다. Reduction 과정은 10mM DTT/100mM ABC 100 μl를 첨가하여 56°C에서 1시간 반응을 시켜 원심분리 후 상등액을 버렸다. 55mM IAA/100mM ABC 100 μl를 첨 가하여 암조건에서 40분 반응을 시킨 다음 원심분리 후 상 등액을 버렸다. 3차 증류수를 IAA의 2~3배를 첨가하고 섞 은 뒤 원심분리하여 상등액을 버리는 과정을 2번 반복하였 다. 100% ACN 100 μl를 첨가하여 5분 동안 섞은 뒤 원심 분리하여 상등액을 버린 다음 다시 원심분리건조기에서 10 분간 건조시켰다. 7~8μl(0.1μg/μl) trypsin enzyme를 첨가 시켰다. 양이 적을 때에는 50mM ABC 40 μl를 더 첨가시 켰다. 37°C에서 12~17시간 놓아두었다. 반응이 끝난 e-tube는 원심분리 후 상등액을 500 μl tube에 넣고 gel이 든 e-tube에 는 50% ACN/5% TFA 50 μl를 첨가하였다. 25°C 실온에서 40분간 놓아두고 원심분리하여 500 μl tube에 넣었다. 다시 gel이 든 e-tube에 100% ACN 50 μl를 첨가하여 원심분리 후 상등액을 500 μl tube에 넣은 후 원심분리건조기에서 완 전히 건조하였다. CBB 염색의 경우는 30% MeOH 200 μl 를 첨가하여 세척하고 50% Acetonitrile/10mA Ammonium bicarbonate 200 μl를 첨가하여 탈색하며 trypsin enzyme을 15 μl를 넣었다. In-gel digestion 후 건조된 시료에 0.5% trifluoroacetic acid (TFA) 10 μl을 가하여 섞은 뒤 원심분 리를 하였다. Zip tip C18 (Millipore, Bedford, MA, USA)을 wetting solution (50% ACN/0.5% TFA/DW)으로 5번 세척 한 후 0.5% TFA/DW로 10번 세척하였다. Pipette을 시료가 녹아있는 e-tube에 넣고 10번 상하로 통과시켰다. 다시 0.5% TFA/DW로 10번 세척하였다. 50% ACN/0.5% TFA/matrix/ DW 4 μl를 취하여 plate에 1 μl씩 옮긴 후 건조시켜 단백 질 동정을 하였다. 단백질 동정에는 대전 기초과학연구소의 Voyager-DE STR MALDI-TOF mass spectrometry를 사용 하였으며 instrument type는 MALDI-TOF-TOF로 하였다.
결과 및 고찰
이차원전기영동을 이용한 개화 후 시기별 단백질 발현 양 상 비교
콩 종실의 개화 후 1주 간격으로 등숙기 동안에 단백질 발현 양상을 비교분석하기 위하여 이차원전기영동을 이용 하였다. 성숙기간 동안은 콩 종자 건물중의 41%는 저장 단 백질이다(Hill and Breidenbach, 1974). 일반적으로 두 가지 종실 저장 단백질은 glycinin과 β-conglycinin이다(Roberts and Briggs, 1965; Hill and Breidenbach, 1974; Thanh and Shibasaki, 1976). 개화 후 1주에서 6주까지 단백질 발현 양 상을 비교하여 보면(Fig. 1), 전반적인 단백질 발현 양상은 등숙이 진행됨에 따라 특별한 차이를 보이지 않았다. 하지 만 단백질 발현 정도는 현저하게 증가하는 것을 확인하였 다. 개화 후 1주에는 단백질 발현 정도가 다른 시기에 비하 여 현저하게 낮았으며 기본적으로 개화 후 2주부터 단백질 발현 정도가 증가하는 것을 알 수 있었다. 개화 후 3주부터 는 확연하게 증가하는 것을 알 수 있었다. 하지만 산성, 중 성, 그리고 염기성부분의 저분자부터 고분자까지 3부분으 로 나누어 단백질 발현 정도를 비교한 결과 등숙이 진행됨 에 따라 모든 단백질의 발현 정도가 점진적으로 증가되지 않는 것을 알 수 있었다. 즉, 등숙이 진행됨에 따라 단백질 의 발현 정도에 단백질 특성(분자량과 수소 이온 농도 지 수)에 따라 차이가 있다는 것이다(Fig. 2). 산성부근의 고분 자부분에서는 단백질 발현 정도가 개화 후 1주부터 3주까 지는 현저하게 증가하다가 개화 후 4주부터는 급격하게 감 소하여 6주에는 1주와 유사한 발현 정도를 보였다. 중성부 근의 30.0-45.0 kDa부분에서는 개화 후 1주부터 5주까지 꾸준히 증가하다가 6주에 다소 단백질 발현 정도가 감소되 었다. 염기성부근의 저분자부분에서는 개화 후 1주부터 6주 까지 연속적으로 단백질 발현 정도가 증가하였다. 등숙기별 단백질 발현 양상을 좀 더 면밀히 관찰하기 위하여 이미지 분석을 통하여 단백질 발현 정도를 수치화하였다.


이미지분석을 통한 개화 후 시기별 단백질 발현 비교 분석
이차원전기영동의 결과를 이용하여 단백질 발현 정도(protein normal volume)를 수치화하기 위하여 20개의 단백질 spot 들을 임의로 산성부분에서 염기성부분까지 그리고 고분자 부분에서 저분자부분까지 선발하였다(Fig. 3). 개화 후 1주 와 2주에서는 단백질 발현 정도가 낮아 확연히 증가하는 3 주를 기준으로 4, 5, 그리고 6주와 비교하여 각각의 단백질 spot의 발현 정도의 증감을 비교하였다(Table 1). 개화 후 3 주부터 6주까지 20개의 단백질 spot들의 평균 단백질 발현 량은 순차적으로 0.48, 0.66, 0.59, 그리고 0.57이었다. 즉, 개화 후 3주를 기준으로 4주에는 대부분의 단백질 spot들의 단백질 발현량이 증가한 반면에, 5주에는 다소 단백질 발현 량이 감소한 단백질 spot들이 있었으며 그 현상은 5주보다 6주에서 다소 심하였다. 결과적으로, 개화 후 4주에는 3주 에 비하여 18개의 단백질 spot들의 단백질 발현량이 증가하 였으며 단지 2개의 단백질 spot들의 단백질 발현량이 감소 하였다. 개화 후 5주에는 3주에 비하여 13개의 단백질 spot 들의 단백질량이 증가하고 7개의 단백질 spot들의 단백질량 이 감소하였다. 개화 후 6주에는 11개의 단백질 spot들의 단백질량이 증가하고 9개의 단백질 spot들의 단백질량이 감 소하였다. 단백질 발현 정도의 양상을 보면 5가지 조합을 나타내는 것으로 확인되었다. 단백질 발현량 양상의 조합을 비교하여 보면, 개화 후 4, 5, 그리고 6주에 따른 증감은 첫 번째 4주(+), 5주(-), 그리고 6주(-), 두 번째는 + - +, 세 번 째는 - - -, 네 번째는 + + +, 다섯 번째는 + + -의 조합을 보였다. 각각의 조합에서의 단백질 spot들의 수를 보면 순 차적으로 4 (+--), 1 (+-+), 2 (---), 9 (+++), 그리고 4 (++-) 이었다. 대부분의 단백질 spot들은 점차적으로 개화 후 6주 까지 단백질 발현량이 증가하였으며, 2개의 단백질 spot들 만의 발현량이 꾸준히 감소하였다. 또한, 특이하게도 1개의 단백질 spot의 발현량은 개화 후 4주에 증가하다 5주에 감 소하는 경향을 보였으나 다시 6주에 증가하는 것을 확인 할 수 있었다.
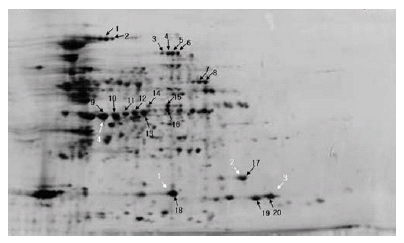
Fig. 3.
Expression of protein in 3 weeks after flowering as basic standard for comparison on protein expression in each week after flowering. White arrows indicate numbers of protein used for MALDI-TOF. Black arrows indicate numbers of protein used for image analysis.Table 1.
Relative protein intensity of differentially expressed proteins from different weeks after flowering in Taekwang.
질량분석을 이용한 단백질 분석
이미지 분석을 통한 결과를 바탕으로 세 번째(---), 네 번 째(+++), 그리고 다섯 번째(++-) 조합에서 총 4개의 spot들 을 임의 선발하여 단백질 동정을 하였다(Fig. 3). 단백질을 동정한 결과(Table 2) spot1은 아직 알려지지 않은 unknown protein으로 동정되었으며 spot3 그리고 4는 저장단백질인 Glycinin A1aBx로 동정되었다. 또한, spot2는 p24 olesin isoform A로 동정되었다. Oleosin은 oil body에 존재하는 기내 번역의 산물로 합성된다. Oleosin의 전사와 동시에 일 어나는 번역과정을 결정하는 것은 특정한 microsomal membrane 이다. 기내 번역반응은 고정된 soybean oil body에서만 이루 어지거나 또는 soybean oil body와 microsome에서 이루어 진다(Loer and Herman, 1993). 식물의 oil-body의 주된 구 성성분은 저장단백질과 막 단백질 그리고 저장지질과 막 지 질이다. Oil body 단백질은 지질단층까지 침투할 수 있으며 지질 상부 그룹과 결합한다. 또한 막 지질 구성은 oil body 와 단백질 결합에 영향을 줄 수 있다(Katavic et al., 2006). 향후, 다른 품종들을 이용하여 품종에 따라 등숙기 동안 단 백질 발현 양상을 비교함으로써 발현되는 단백질에 차이가 있는지 확인할 필요가 있을 것이며 좀 더 면밀한 연구를 위 하여 배와 배유를 나누어 분석함으로써 어떤 단백질이 배와 배유에서 어떻게 등숙이 진행됨에 따라 단백질 발현 양상이 달라지는지 분석할 필요가 있을 것으로 사료된다.
Table 2.
List of identified protein spots from two-dimensional gel using MALDI-TOF-TOF.
적 요
본 연구는 국내 육성 품종인 태광콩의 등숙기에 따른 단 백질 발현 양상을 비교함으로써 등숙기 단백질 발현의 차이 에 대한 기초자료를 얻고자 수행하였다. 동한 개화 후 종실 의 등숙이 진행됨에 따라서 단백질 발현 양상이 세가지 경 향으로 나뉘어 지는 것을 확인하였다. 첫 번째는 등숙이 진 행됨에 따라서 단백질 발현 정도가 증가하다가 감소되며, 두 번째는 증가와 감소의 시기가 성숙기에 이루어지며, 세 번째는 등숙기부터 성숙기까지 점진적으로 증가하는 것이 다. 이러한 현상은 단백질의 기능에 따라 달라지는 것으로 사료된다. 등숙 초기에는 등숙에 필요한 단백질의 발현이 증가할 것이며 등숙 후기에는 저장단백질의 발현이 증가할 것으로 사료된다. 따라서 향후 좀 더 많은 수의 단백질 spot 들을 동정하여 어떤 기능을 가진 단백질이 등숙기에 따라 단백질의 발현 양상이 달라지는지는 좀 더 면밀히 관찰할 필요성이 있다고 사료된다.
