

서 론
재료 및 방법
공시재료
핵심집단 생육 및 엽록소 함량 측정
DNA 추출 및 유전형 데이터 확보
전장유전체연관분석(GWAS, Genome-Wide Association Study)
가뭄스트레스 연관 SNP 발굴 및 후보 유전자 발굴
결과 및 고찰
표현형 데이터 분석
SNPchip을 통한 유전형 데이터 정리
GWAS 수행결과 및 선발된 SNP에 대한 검정
가뭄 스트레스 연관 유전자 후보군 발굴
적 요
서 론
밀(Triticum aestivum L., 2n = 6x = 42, AABBDD)은 세계 인구의 약 40%가 주식으로 삼고 있는 3대 주요 식량 작물이다(Nam & Hahn, 2000; Oh et al., 2019; Pang et al., 2020; Zhao et al., 2018). 전 세계적으로 매년 약 7억 6,050만 톤의 밀이 생산되며 재배 면적은 약 2억 1,700만 ha에 해당한다(Ray et al., 2013). 밀은 벼나 옥수수보다 영양성이 우수하여 인간이 섭취하는 영양분 중 35%의 칼로리를 밀에서 흡수하고 있다(Ali et al., 2020). 밀알의 영양성분은 대략 탄수화물 70~74%, 단백질 10~14%, 지방 1.9~ 2.3%로 구성되며 티아민, 리보플라빈 등의 비타민류도 풍부하여 영양적 가치가 우수하며, 미네랄 등 유익한 성분이 많이 함유되어 있다(Ali et al., 2020).
전 세계적인 기후 변화는 작물 재배 환경에 영향을 미치고 있으며 밀 재배지에서는 환경 스트레스에 대한 작물의 피해를 최소화하는 것이 수량을 유지할 수 있는 중요한 요인으로 사료되고 있다(Guedira et al., 2016). 그 중 가뭄 스트레스는 작물에 미치는 피해와 영향력이 막대하여 미국 해양기상청에서는 가뭄을 20세기 최대 자연재해 중 상위 5위 안에 드는 기상재해라고 규정하고 있다(Bae et al., 2015). 2021년에는 미국과 브라질의 극심한 가뭄으로 인하여 세계 식량 가격이 최근 10년 만에 가장 높은 수준으로 상승하고 곡물가격의 안정성이 떨어져 전세계의 식량안보에 위협이 되고 있다(Yang et al., 2023). 국내의 경우에는 2015년 이후에 지역별로 국지적인 가뭄이 매년 발생하고 있고, 가뭄의 발생빈도와 강도가 증가하는 추세이며 특히, 우리나라의 기후 특성으로는 4~5월의 가뭄과 6~7월의 장마기가 작물에 영향을 미치는 것으로 보고되어 있다(Park et al., 2020; Jeon et al., 2021). 특히, 영양생장시기에 가뭄의 영향을 받으면 토양 수분의 부족으로 인해 밀의 생육이 저하되어 종실 수량이 크게 저하된다고 한다(Yang et al., 2023). 따라서 작물의 수확량 개선 및 가공적성에 영향을 미치는 것을 최소화하기 위하여 영양생장기와 생식생장기에 가뭄 저항성이 높은 밀 자원을 탐색하고 가뭄 저항성 연관 유전자를 발굴하여 육종에 활용하는 것은 매우 중요한 연구라고 할 수 있다(Sallam et al., 2019; Yang et al., 2023).
가뭄 스트레스는 작물의 엽록소 함량을 감소시키고, 광합성 메커니즘을 파괴하며, 성장을 억제하고, 궁극적으로 수량을 크게 저하시키며(Ali et al., 2020; Sharifi & Mohammadkhani, 2016; Yang et al., 2023), 특히, 지엽의 엽록소함량으로 가뭄의 영향을 판단할 수 있다고 한다(Ali et al., 2020; Qian et al., 2021). 식물에서 엽록소의 함량은 온도, 물, 질병 및 기타 스트레스를 포함한 다양한 요인의 영향을 받고 있으며(Hörtensteiner & Kräutler, 2011). 특히 수분 부족은 작물에서 잎의 엽록소 합성에 영향을 미치고, 엽록소의 분해를 가속하여 빛 에너지의 흡수 능력을 감소되어 광합성을 억제하게 된다(Hörtensteiner & Kräutler, 2011). 따라서, 잎에서의 엽록소 함량은 가뭄 스트레스 측정지표로 사용되며 가뭄 저항성 연관 유전자의 탐색에 활용되고 있다(El Gataa et al., 2022; Devate et al., 2022). Javed et al. (2022)는 엘리트 품종의 성숙기에 수량, 엽록소 함량, 단백질 함량을 통하여 가뭄 스트레스의 저항성을 측정하였다.
최근 유전체학 분야의 발전으로 많은 작물에서 유전체 기반 육종 연구가 진행되고 있으나 16G의 유전체를 가진 거대배수성 작물인 밀에서는 완벽한 전체 유전체의 해독이 이루어지지 않았으며 유전체 기반 연구에 어려움이 있어 밀 유전자원의 대표성을 가지고 있는 한국형 밀 핵심집단을 이용한 연구가 필요하다(Kang et al., 2023; Lee et al., 2023; Paux et al., 2006). 핵심집단(Core collection)은 모집단의 유전적 다양성을 대표하도록 최소한의 자원으로 구성된 집단이며, 국내에서는 2020년에 한국형 밀핵심집단을 구축한 사례가 있다(Frankel & Brown 1984; Min et al., 2021). 또한, 유전체 기반 육종에는 염기서열 분석 유전형 분석 기법인 GBS (Genotyping-by-sequencing)가 많이 사용되었으며 밀의 유전체 전체를 커버하는 수많은 단일 뉴클레오티드 다형성(SNP) 마커가 발굴되고 있다(Hussain et al., 2017; Sallam et al., 2019). 그러나 GBS에 의해 생성된 각 SNP에 대한 정확한 위치와 염색체상의 위치는 이질 6배체인 밀에서는 확인하기 어려운 한계가 있다. 최근에는 기존에 보고된 SNP를 이용하여 집단내에서 전장유전체연관분석(Genome-Wide Association Study, GWAS) 및 양적 형질 유전자좌(QTL) mapping을 가뭄 저항성의 유전적 특성 분석에 이용하고 있다(Sallam et al., 2019). 가뭄과 같은 특정 환경 조건에서 형질과 연관된 유전체 지역을 탐색하려면 QTL과 같은 관련 유전체 영역의 조사가 필요하며, GWAS는 연관 불균형(Linkage Disequilibrium)을 기반으로 탐색하고자 하는 형질과 높게 연관된 유전자 또는 QTL을 식별하는 데 효과적인 방법 중 하나라고 알려져 있다(Devate et al., 2022). 또한, 후보 유전자 예측을 위해 GWAS를 활용하여, 밀에서도 성숙기 단계에서 가뭄 스트레스 저항성과 연관된 유전체 영역을 탐색한 사례가 있고 Axiom사의 35K SNP genotyping array를 활용하여 엽록소와 연관된 GWAS를 분석하여 가뭄 저항성 SNP를 발굴하기도 하였다(Devate et al., 2022; El Gataa et al., 2022; Reddy et al., 2023).
해당 연구는 금강밀을 포함한 국내 품종 5개가 포함된 핵심집단을 활용하여 유묘기, 성숙기에 가뭄처리를 받은 밀 식물체의 엽록소 함량을 측정하고 대조구와 비교하였다. 이를 기반으로 가뭄 스트레스에 대한 저항성 계통 탐색 및GWAS 분석을 통해 SNP를 탐색하고 후보유전자군 및 마커로서의 활용 가능성을 밝히고자 한다.
재료 및 방법
공시재료
본 연구를 진행하기 위하여 국내 대표 장려품종인 금강밀을 포함한 130점의 국내 밀과 멕시코의 108점, 미국의 52점, 중국의 33점, 일본의 10점과 다른 기타국가(46개국)의 을 276점을 포함한 한국형 밀 핵심집단 609점을 농촌진흥청 농업유전자원센터에서 분양받았다.
핵심집단 생육 및 엽록소 함량 측정
분양받은 밀 핵심집단 종자는 공주대학교 연구포장(40 m × 32 m)에서 40 m인 가로 1줄에 핵심집단 40계통씩을 각 계통마다 1 m씩 간격을 두고 2021년 10월 18일에 파종하였다. 567계통의 유묘기 식물체는 32구 규격의 모종트레이에서 파종 후 12주동안 정상생육을 진행한 후에 건조처리를 하였으며 567계통의 성숙기 식물체는 출수시기(2022년 4월 20일) 직전에 가로 55.8 cm, 세로 19.6 cm, 높이 15.8 cm인 초화박스에 옮겨 심어 비가림 온실에서 출수 및 등숙을 진행하였다. 모종트레이로 옮겨 심은 핵심집단에 대하여 유묘기에는 3일 무관수처리 후 엽록소 측정을 바로 진행하였고, 성숙기에는 Baek et al. (2009)를 응용하여 3일 무관수-1일 관수(계통 당 50 mL)를 1번의 처리구로 하여 무관수 처리(유닛1) 다음날 엽록소 함량을 측정하였다. 무관수 처리 2반복(유닛2), 3반복(유닛3)을 위한 무관수의 시작은 엽록소 함량 측정 다음날부터 시작하였다. 각 유닛에 적용된 계통 당 50 mL의 관수는 식물 생장을 위한 최소 수분이며 농촌진흥청의 토양수분 장력 측정 시 사용되는 기준을 적용하였다(RDA, 2012). 각 대조구와 처리구에서 SPAD-502(KONICA MINOLTA, Tokyo, Japan)를 활용하여 유묘기에서는 제일 마지막에 나온 잎을, 성숙기에는 지엽에서 각 3반복으로 엽록소를 측정하였다. 유묘기의 엽록소 측정은 전체 집단에 대한 1회의 측정이므로 각 반복에 대한 평균값을 구하였으며 대조구에 대한 처리구의 비율도 계산하였다. 성숙기는 각 유닛별 측정된 평균값과 유닛 전체에 대한 평균값을 각각 계산하였으며 마찬가지로 대조구에 대한 처리구의 비율도 계산하였다. 유묘기 식물의 경우 모종트레이의 크기가 작고 높은 재식밀도로 인한 식물 스트레스를 고려하여 토양수분함량을 측정하지 않았으나 성숙기에서는 HMM-200 pro (Han young systems, Seoul, Korea) 토양수분계를 사용하여 각 유닛마다 토양수분함수율(SWC, Soil water content)을 측정하였다. 핵심집단 성숙기의 가뭄 스트레스 지수(DTI, Drought Tolerant Index)는 Bonfim-Silva et al. (2015); Huh & Lee (2022)을 적용하여 토양수분함수율에 대한 엽록소의 함량(chlorophyll content)으로 정의하여 아래의 식으로 도출하였다.
GWAS에 사용된 표현형데이터는 각 유닛 별 DTI를 활용하여 가뭄처리 대비 대조구와의 차이와 비율을 작성하였다.
DNA 추출 및 유전형 데이터 확보
패트리 디시에서 자란 10일 된 어린 잎을 채취하여 액화질소로 곱게 마쇄한 후 Genomic DNA Prep Kit (Biofact, Daejeon, Korea)를 활용하여 Genomic DNA를 추출하였다. 추출된 DNA 농도는 20 ng/ul으로 조절하여 분석할 때까지 -20℃에 보관하였다. 유전형 데이터는 한국형 밀 핵심집단 전체 609점 중 DNA QC (DQC of 0.82, QC call rate 97%)와 국내 생육조건에 정상적 재배가 가능한 계통으로 선발하여 최종 567점을 이용하여 분석하였다. 밀 핵심집단 내 567점의 DNA를 35,143개의 SNP마커가 포함된 Axiom회사의 상용 SNP array인 BreedWheat 35K SNP chip (Thermo Fisher Scientific, MA, USA)을 이용하여 유전형 데이터를 확보하였고 Axiom Analysis Suite 5.2 소프트웨어(Thermo Fisher Scientific, MA, USA)로 분석하였다(https://www.cerealsdb.uk.net/cerealgenomics/cerealsdb/axiom_download.php).
분석된 SNP chip 데이터 중에서 minor allele frequency(MAF)가 5% 미만이면서 missing value가 10% 초과되는 SNP들을 적합하지 않은 SNP로 판단하여 제거하였으며, Beagle v5.2와 Tassel v5.2.57 같은 생물정보학 도구를 활용하여 SNP chip 필터링을 진행하였다(Bradbury et al., 2007; Browning et al., 2018).
전장유전체연관분석(GWAS, Genome-Wide Association Study)
수집한 표현형 데이터인 엽록소 함량차이(가뭄 처리구의 엽록소 함량 – 대조구의 엽록소 함량)과 비율(가뭄 처리구의 엽록소 함량/대조구의 엽록소 함량)에 대하여 SNP chip data를 GAPIT version3의 R프로그램을 이용하여 각각 GWAS를 수행하였다. GWAS 분석에 사용된 모델은 mixed linear model (MLM), Fixed and random model circulating probability unification (FarmCPU), Bayesian-information and linkage-disequilibrium iteratively nested keyway (BLINK) 3가지 모델로 수행하였으며 각 모델별로 도출된 결과를 서로 비교분석 하였다.
가뭄스트레스 연관 SNP 발굴 및 후보 유전자 발굴
GWAS 결과로 선발된 SNP들에 대하여 표현형데이터로 상하위 5계통들의 유전형을 조사하여 SNP 마커로 활용이 가능한지 판별하였다. GAPIT3의 cut off 기준을 토대로 Bonferroni correction (0.05)를 활용한 SNP 마커수(22,775)의 계산으로 나온 p-value에 log 값을 취하여 LOD를 도출하였으며 LOD가 5.65 이상인 SNP를 선발하였다. 또한, 선발된 SNP에 대해 567계통의 haplotype 분석을 통하여, 마커로서의 활용 가능성을 탐색하였다. 후보유전자군의 경우 IWGSC v1.0 genome sequence를 참조하여 SNP를 기준으로 ±250 kb범위를 탐색하였다. 선발 SNP 기준 ±250 kb에서 후보유전자를 선발한 이유는 국내품종을 가지고 GWAS를 수행한 선행연구에서 밀염색체의 연관불균형(LD, Linkage Disequilibrium)값을 최소 74.7 kb, 최대 393 kb로 추정하여 보고되었기 때문에, 대략적인 평균값인 250 kb에서 후보 유전자를 추정하여 활용하였고 이러한 방식은 밀 GWAS 분석에 활용되고 있다(Jung et al. 2021; Li et al., 2019; Zhao et al., 2020; Zhu et al., 2021).
결과 및 고찰
표현형 데이터 분석
밀 핵심집단 계통 중에서 국내 생육조건에 맞춰 정상적으로 생육이 가능한 567계통을 영양생장기인 유묘기와 생식생장기인 성숙기에 대하여 엽록소를 측정하였다. 유묘기의 무처리 조건에서의 엽록소 함량은 최소 21.10 mg/m2에서 최대 45.83 mg/m2까지 나타났으며, 표준편차는 3.506, 가뭄 스트레스 조건에서는 최소 0.57 mg/m2에서 최대 37.33 mg/m2로 표준편차는 6.924로 측정되었다. 성숙기의 엽록소 함량은 무처리에서 최소 1.41 mg/m2에서 최대 50.42 mg/m2로 표준편차는 1.171이며, 가뭄 스트레스 조건에서 최소 3.24 mg/m2에서 최대 43.04 mg/m2로 표준편차는 1.426으로 측정되었다. 유묘기에서의 가뭄스트레스에 대한 엽록소 함량의 분포도는 정규분포를 보이고 있으며 정상적인 표현형 측정이 진행되었다고 판단된다(Fig. 1). 성숙기에서의 무처리에서의 DTI 지수는 최소 0.27에서 최대 10.36의 스트레스 지수가 나왔으며 가뭄 스트레스 조건에서의 DTI 지수는 최소 0.73에서 15.62의 값이 나타났다. 또한, 엽록소 함량만으로는 저항성 정도를 판단하기 어렵기 때문에 성숙기 무처리 조건에서의 엽록소 함량에 대한 가뭄 스트레스의 엽록소 함량의 비율(처리구의 엽록소 함량/대조구의 엽록소 함량)을 분석하였다(Table 1, Fig. 1C, Fig. 2C, F, I, L). 유묘기의 건조처리와 무처리의 엽록소 비율은 평균 0.534, 최솟값 0.021, 최댓값은 1.157로 나타났으며 전체적으로 정규분포를 나타내고 있다(Fig. 1C). 반면 성숙기의 엽록소 비율은 시간이 지남에 따라 누적되므로 값을 나타냈을 때, 무관수 처리인 유닛1(Fig. 2A, B, C)은 평균1.121, 최솟값0.039, 최댓값9.410으로 무관수 처리 2반복인 유닛2(Fig. 2D, E, F)는 평균1.734 최솟값 0.012, 최댓값 27.090으로, 무관수 처리3반복인 유닛3(Fig. 2G, H, I)은 평균 3.600, 최솟값 0.148, 최댓값 222.330으로 나타났다. 각 유닛별 DTI의 평균값은 유닛1보다 유닛3으로 갈수록 커지는 경향이 보이는데 이는 성숙기의 대조구에서 엽록소 함량이 유닛1에 비하여 유닛3에서 감소하였기 때문이다. 이 결과는 가뭄 스트레스 처리 기간 동안(1처리구당 4일, 총 3처리구는 12일) 성숙기 핵심집단 밀의 노화가 진행되어 대조구의 엽록소가 감소하였으나 처리구는 가뭄 스트레스에 저항하기 위하여 엽록소 함량의 감소 폭이 상대적으로 적어 유닛3의 엽록소 비율이 높아진 것으로 사료된다. Wasaya et al. (2021)에 따르면, 가뭄 스트레스에 저항성을 가지고 있는 계통인 Galaxy-2013에서 엽록소 함량이 대조구와 비교하여 더 높았고 잎의 녹체의 유지(staying green)가 가장 길게 관찰되었다. Javed et al. (2022)은 14개의 엘리트집단에 건조스트레스를 처리하고 엽록소 함량을 측정하였으며, 건조스트레스를 처리한 집단이 정상적인 관수처리를 진행한 집단보다 엽록소 함량이 증가된 경향을 보였다. 이 결과는 가뭄 스트레스에 대응하기 위한 식물의 반응으로 유닛3의 실험결과에서 처리구의 DTI가 대조구보다 높은 결과와 유사한 양상을 보여준다(Table 1). 성숙기에서의 유닛 전체의 엽록소 비율 평균값이 1.317이며, 최솟값은 0.216, 최댓값은 9.890으로 유묘기에서의 비율 보다 차이가 크게 나타났다(Fig. 1C, Fig. 2J, K, I). 대조구에 대한 가뭄 스트레스 처리구의 비율을 기준으로 핵심집단 중 567점의 1%에 해당하는 상하위 5계통에 대한 값을 유묘기와 성숙기의 엽록소의 차이, 비율별로 나타내었다(Table 1). 유묘기 시기에서는 cc31, cc1884, cc26, cc507, cc118계통들의 엽록소 비율이 각각1.157, 1.096, 1.089, 1.072, 1.030, 평균 1.089로 높게 측정되었으며, cc936, cc794, cc585, cc1010, cc856계통들의 엽록소 비율이 각각 0.096, 0.079, 0.065, 0.050, 0.021, 평균 0.062로 저조하였다(Table 1). 성숙기에서 처리구와 대조구간 엽록소 함량차이에서는 상위 계통 평균값은 7.173, 하위 계통의 평균값은 -3.622로 나타났다. 비율 부분에서는 cc113, cc1234, cc694, cc512, cc110 계통의 엽록소 비율이 각각 9.890, 8.199, 6.587, 6.096, 5.372, 평균 7.229로 높게 측정되었으며, cc31, cc484, cc4, cc1796, cc479계통의 엽록소 비율이 각각 0.510, 0.406, 0.388, 0.388, 0.358, 0.216, 평균 0.375로 낮게 나타났다(Table 1). 유묘기에서는 생육우수자원 후보군인 상위 5계통의 경우 엽록소 비율이 높은 계통에서의 평균값이 1.089로 나타났으나, 성숙기에서는 상위 5계통의 경우 엽록소 비율이 높은 계통의 평균값이 7.229이다. 두 생육시기에서의 엽록소 함량과 비율값을 조사한 결과 상위 5계통들 총 20계통 중에서 cc1884와 cc694는 엽록소 함량과 비율에서 유묘기와 성숙기에 각각 중복되었으며, cc31은 유묘기 비율에서는 상위 계통이였으나 성숙기 비율에서는 하위 계통으로 평가되어 제외하였다. 이를 통하여 최종 17계통을 가뭄저항성 자원으로 선발하였다. 가뭄 스트레스의 경우 엽록소 함량의 비율이 유묘기에 대비해서 성숙기에 더 큰 영향을 미치는 것으로 예측되며, 이는 유묘기의 엽록소 보다는 성숙기의 노화된 잎에서 더 많은 영향을 받았을 것으로 예상된다. 또한, 성숙기에서는 표현형 데이터의 분포도가 전반적으로 오른쪽꼬리분포(왜도, Skewness)로 나타났으며, 비율값으로는 0~2점대의 값을 가진 핵심집단 계통들이 540점으로, 이는 전체 핵심집단의 약 80%에 속한다. 그래프를 통해 성숙기에서의 비율값을 분석한 결과, 가뭄 스트레스 저항성이 강한 계통들이 핵심집단 내에서 극소수임을 확인할 수 있었다. Javed et al. (2022)에 따르면, 가뭄 스트레스 조건에서 대조구보다 처리구의 엽록소 함량이 높아지는 경향이 나타났으며, 유사한 반응이 가뭄 처리 핵심집단에서도 나타난 것으로 사료된다(Fig. 2).
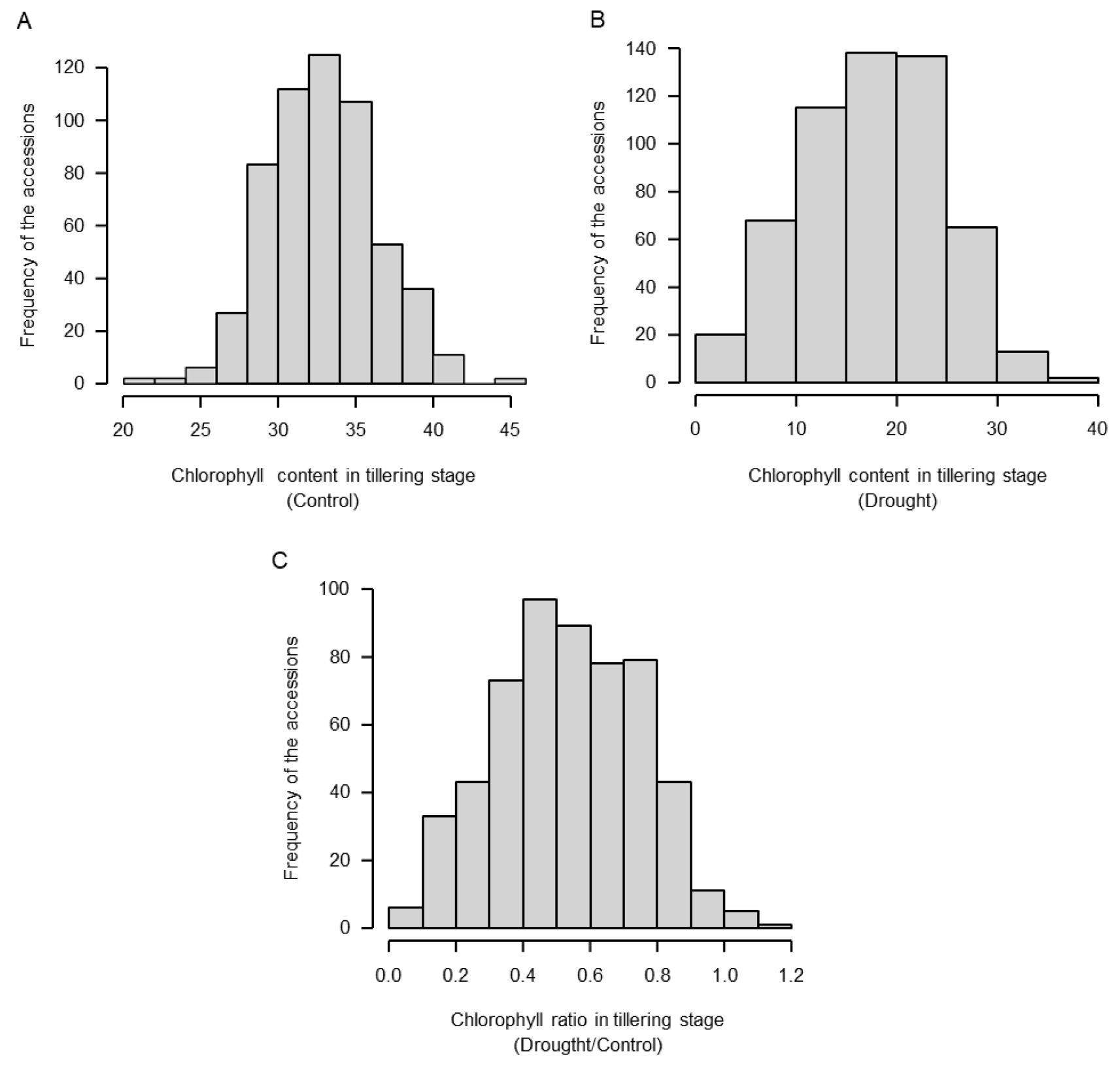
Table 1.
Chlorophyll data and drought stress index (DTI) of the top five and bottom five accessions during the tillering stage and ripening stages under drought stress.
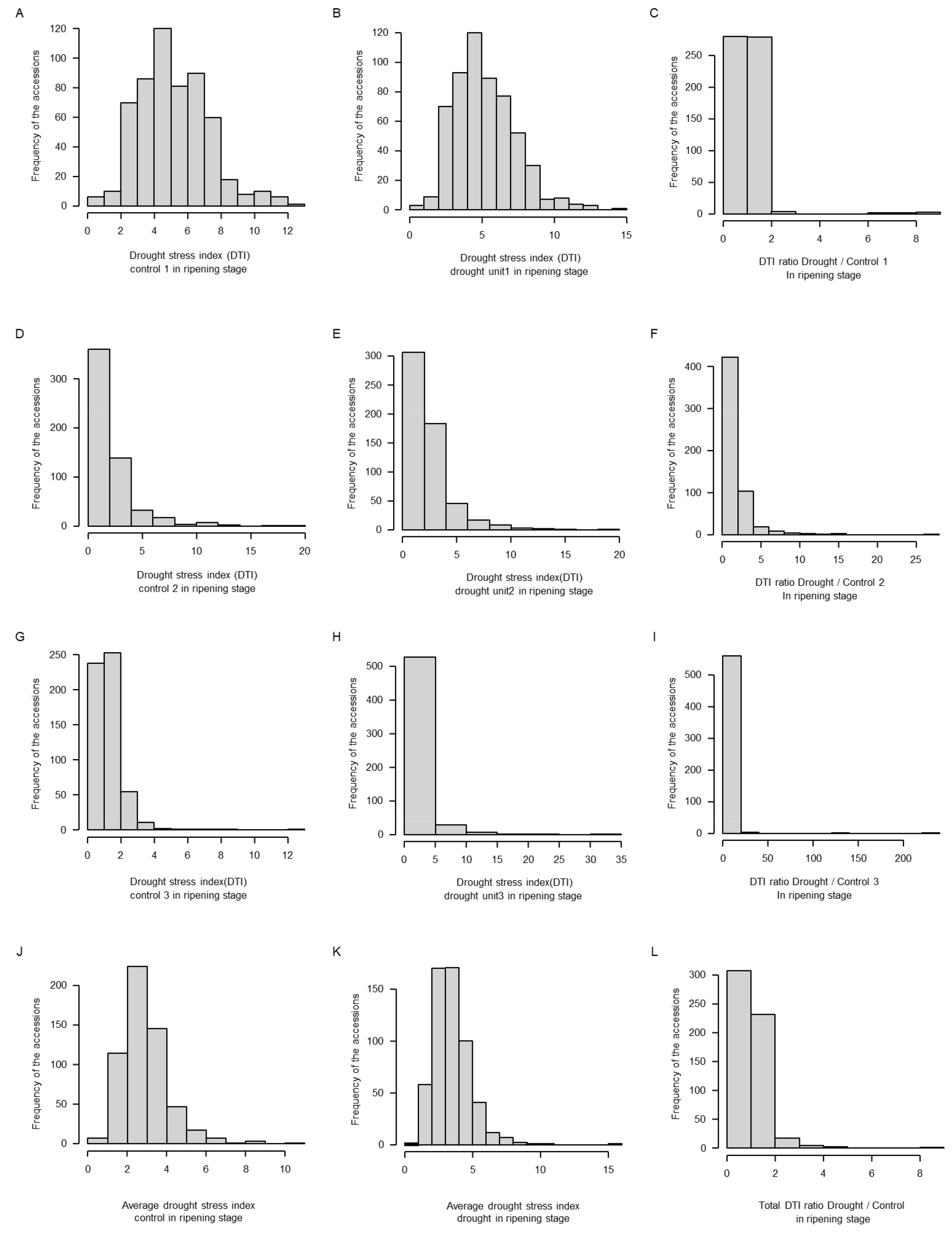
Fig. 2.
Frequency distribution of accessions to wheat core collection DTI during the ripening stage under non-treatment, drought stress treatment, and average (A: control 1; B: drought unit 1; C: ratio D/C1; D: control 2; E: drought unit 2; F: ratio D/C2; G: control 3; H: drought unit3; I: ratio D/C3; J: total control average; K: total drought average; L: total ratio D/C).
SNPchip을 통한 유전형 데이터 정리
핵심집단에 대한 SNP chip 분석을 통해서 전체 35,143마커 중에서 위치가 알려지지 않은 마커 및 MAF (Minor allele frequency) 5% 미만을 제거한 후 총 22,775개의 SNP 마커를 기반으로 추후 분석을 수행하였다. 이러한 SNP들은 모든 염색체에 분산되어 있는데, A 게놈에는 7,065 (31.1%), B 게놈에는 8,480 (37.2%), D 게놈에는 7,230 (31.7%)개의 마커가 존재하며, B 게놈이 다른 게놈보다 5.5~6.1% 더 많은 SNP 마커를 포함하고 있다는 것으로 나타났다.
GWAS 수행결과 및 선발된 SNP에 대한 검정
GWAS의 MLM, FarmCPU, BLINK의 3가지 모델을 이용하여 유묘기 및 성숙기의 표현형 데이터와 연관된 SNP들을 탐색하였다(Fig. 3). 활용한 모델 중 MLM의 경우에는 고정효과와 임의효과를 모두 포함한 혼합선형 모델이며, FarmCPU의 경우에는 pseudo-quantitative trait nucleotide를 활용한 친족관계(kinship matrix)를 계산하여 통계적 검정력이 향상되었으며, BLINK의 경우에는 연관불균형 정보(LD)와 Bayesian information criteria (BIC)함수로 컴퓨팅 시간을 단축하는 특징이 있다(Cebeci et al., 2023). 본 실험에서는 거대 6배체 작물인 밀 유전체에 적합한 모델을 확인하고 선발된 SNP의 재현성을 검증하기 위하여 MLM, FarmCPU, BLINK의 3가지 모델을 동시에 분석하였다. SNP 선발은 MLM, FarmCPU, BLINK 분석 결과에서 LOD가 5.65 이상이 되는 것으로 하였다. 유묘기에서는 3가지 모델에서 공통적으로 발견된 SNP가 없어서 성숙기에서만 선발하였으며 1A, 1B, 5B, 5D, 7D에서 각각 1개의 SNP가 탐색되었다. 이 결과는 Fig. 3에서 맨해튼 플롯과 빨간 박스로 표시하였다.
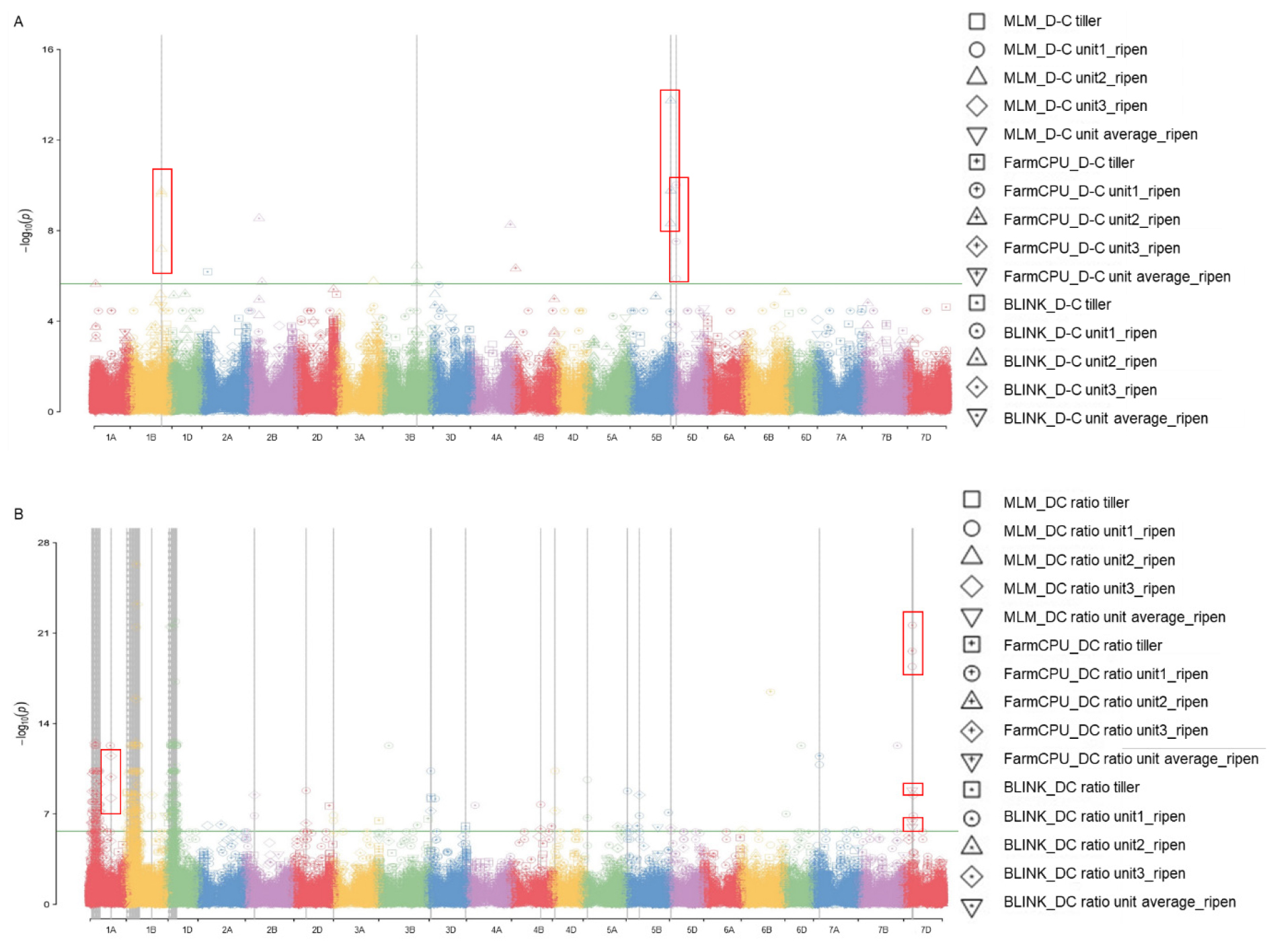
선발된 SNP들은 성숙기에서의 AX-95204454, AX-95238538, AX-95260370, AX-95139321, AX-94434693로 이들에 대하여 상하위 5개 계통의 엽록소 데이터와 유전자형을 분석하였다(Tables 2, 3). 또한 선발된 SNP에 대한 전체 집단에서 표현형과의 연관분석을 진행한 결과, 성숙기에서는 AX-95260370, AX-95139321, 이 2개의 SNP에서는 유의성이 나타나지 않았다(Fig. 4). 이에 따라, AX-95204454, AX-95238538, AX-94434693, 총 3개의 SNP들에 대해 하플로타입(haplotype)분석을 진행하여 마커로서의 가능성을 탐색하였다(Table 4). 전체 핵심집단 내 하플로타입과 상하위 계통 분석 및 Fisher’s least significant difference (LSD)를 활용하여 alpha값을 0.05로 설정 후에 분석을 진행한 결과 유전형에 따른 표현형 값의 유의성이 존재하였다. AX-95204454의 경우에는 동형접합자인 GG가 상위계통, 이형접합자인 AG가 하위계통에서 나타났다(Fig. 4). 또한, 유전형에 따른 표현형이 AX-94434693, AX-95238538와 같이 8계통(AA), 10계통(CC), 9계통(TT)으로 치우쳐져 있지 않다(Table 4). 상하위 5계통에 대한 대조를 통해 유전자형에 따른 표현형의 유의성이 나타났으므로 추후 마커로 활용이 가능할 것으로 사료된다(Tables 2, 3). 하나의 SNP로 양적 형질과 같은 가뭄 스트레스의 저항성을 판단하기에는 다른 요소들도 고려해야 하나 추후 유전자후보군의 염기서열 결과를 참조하여 가뭄 스트레스에 영향을 미치는 관련 유전자를 탐색할 수 있을 것으로 사료된다.
Table 2.
Genotypes of AX-95204454, AX-95238538, and AX-95260370 for the top and bottom five accessions based on the DTI difference values during the ripening stage.
Table 3.
Genotypes of AX-95139321 and AX-94434693 for the top and bottom five accessions based on the DTI ratio values during the ripening stage.
| Accession No. | AX-95139321 | AX-94434693 | |
| Top5 | cc113 | GG | AG |
| cc1234 | AG | GG | |
| cc694 | AG | AG | |
| cc512 | AA | AG | |
| cc110 | GG | AG | |
| Bottom5 | cc31 | AG | GG |
| cc484 | GG | AG | |
| cc4 | AG | GG | |
| cc1796 | AG | GG | |
| cc479 | GG | AG |
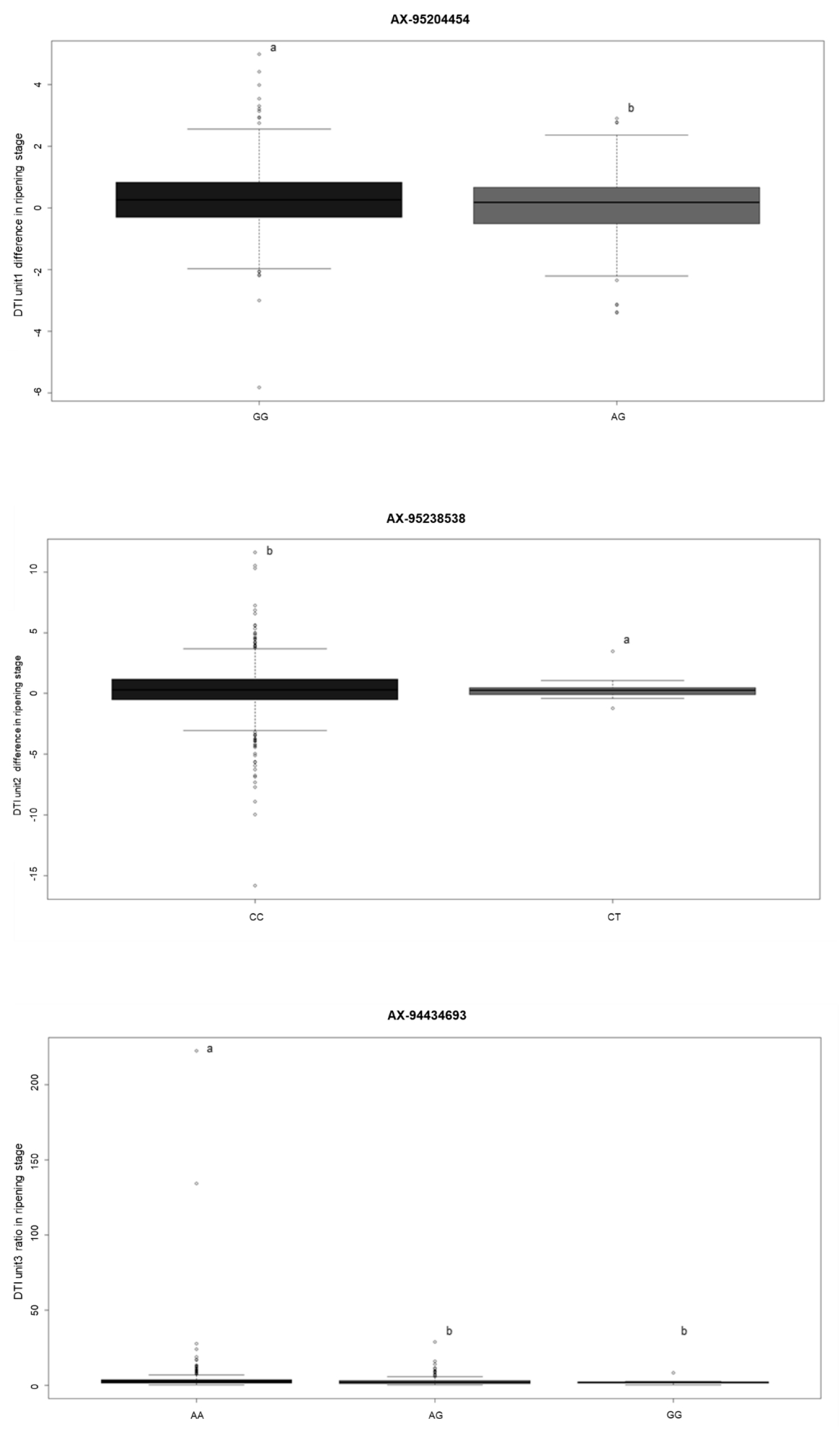
Table 4.
Haplotype variations in AX-95204454, AX-95238538, and AX-94434693. Phenotypic data are presented as mean and standard deviation within the group as DTI (difference, ratio) values.
가뭄 스트레스 연관 유전자 후보군 발굴
성숙기에서의 엽록소 표현형 데이터에서 탐색된 SNP들 중에서 MLM, FarmCPU, BLINK에서 공통적으로 발견된 5개의 SNP들 주변으로 ±250 kb를 탐색한 결과, uncharacterized protein을 제외하고 총 16개의 유전자 후보군을 탐색하였다(Table 5). 3개의 GWAS모델에서 모두 선발된 AX-95139321은 유전자 내에 존재하는 SNP로서, 염기서열이 G또는 A가 치환될 경우, 아미노산 서열이 바뀌는 missense 돌연변이를 유발하게 된다. AX-95260370도 마찬가지로 염기서열의 치환이 아미노산에 영향을 미치는 missense로 나타났으며, AX-95204454는 3’-UTR 영역에, AX-95238538과 AX-94434693은 Intergenic 영역에 속해 있다. 발굴된 유전자 중에서 흥미로운 유전자 후보로는 TraesCS5B02G505500, TraesCS7D02G188000가 있으며 이들은 탐색된 SNP가 치환될 경우 아미노산이 바뀌게 되어 이 유전자의 기능이 상실될 것으로 예상된다. AX-95260370이 속한 TraesCS5B02G505500은 F-box domain으로 F-box 패밀리의 경우 개화, 종실 발달 및 잎의 노화와 호르몬 신호 전달들을 포함한 여러 생물학적 과정에서 단백질 분해에 특정한 역할을 한다는 것이 밝혀졌으며 이전 연구에서 qRT-PCR을 통해 밀의 생장과정에서 일부 F-box 유전자가 영양생장시기 또는 종실의 발달에서 특이적으로 발현된다는 것이 밝혀졌다(Hong et al., 2020). 또다른 missense SNP인 AX-944434693이 속한 TraesCS7D02G188000(aldehyde oxygenase)는 알데히드 탈수소효소(ALDH)로 식물 대사에 관여하고 알데히드 항상성에 기여하여 독성 알데히드를 제거하는 효소 계열로서, ALDH 효소는 효소 반응에서 NADPH와 NADH를 생성하여 산화환원 등가물의 균형을 맞추는 데 기여한다(Zhao et al., 2017). 위와 같이 발굴한 유전자의 직접적인 가뭄 스트레스 저항성 여부는 추가적인 발현 검정을 통하여 분석하여야 한다.
본 연구는 밀 핵심집단의 유묘기와 성숙기에 가뭄 스트레스를 처리하여 엽록소 함량을 측정하고 대조구와의 엽록소 함량의 차이와 비율을 이용하여 연관분석을 진행한 연구이다. 표현형 분석을 통하여 핵심집단 내에서 가뭄 스트레스에 강한 17계통을 선발하였으며 GWAS 연관 분석을 통하여 AX-95204454는 마커로 개발될 가능성이 있는 것으로 확인되었다. 또한 발굴된 가뭄 저항성 후보 유전자는 추가적인 분석을 통하여 내건성 밀 육종에 활용될 수 있을 것이다.
Table 5.
Candidate genes were explored within ± 250 kb of AX-95204454, AX-95238538, AX-95260370, AX-95139321, and AX-94434693. Underlined gene IDs indicate that the identified SNP is located within the specified gene.
적 요
본 연구는 밀의 생육시기 중 유묘기와 성숙기에 가뭄 스트레스를 부여하고 각 시기에 측정한 엽록소 함량을 비교하여 가뭄 저항성 계통과 생육우수자원을 선발하고 GWAS를 통해 가뭄 스트레스와 연관된 SNP및 유용유전자를 탐색한 결과는 다음과 같다.
1.핵심집단에서 가뭄 스트레스에 대하여 저항성 계통으로 선발된 자원은 시기별로 수집한 엽록소의 함량차이값과 비율, DTI를 고려하여 상,하위 5계통에 동시에 속한cc31을 제외한 상위 계통인cc12, cc26, cc102, cc110, cc113, cc118, cc507, cc512, cc576, cc694, cc834, cc845, cc1234, cc1360, cc1792, cc1884, cc1912 총 17계통이 선발되었다. 특히 cc694와 cc1884는 엽록소 함량차이값과 비율에서 모두 선발된 자원이다.
2.전장유전체 연관분석(GWAS) 결과 1A, 1B, 5B, 5D, 7D에서 유의성이 있는 SNP를 발굴하였으며 발굴된 SNP는 AX-95204454, AX-95238538, AX-95260370, AX-95139321, AX-94434693였다.
3.발굴된 SNP를 중심으로 인근의 후보 유전자군을 분석한 결과 TraesCS7D02G188000 (aldehyde oxygenase), TraesCS5B02G505500 (F-box domain)을 발굴하였다.
4.AX-95204454는 SNP 중에서 전체 핵심집단 내 하플로타입 분석과 상하위 5계통에 대한 대조를 통해 유전자형에 따른 표현형의 유의성이 있어 추후 성숙기에서의 엽록소 함량차이(DTI_difference)를 기준으로 가뭄저항성 계통 선발 마커로 활용이 가능할 것으로 사료된다.
