

서 론
재료 및 방법
실험 재료
귀리 가수분해물 및 분말 시료 제조
일반성분 분석
구성 아미노산 분석
단백질 품질 분석
아베난쓰라마이드 함량 분석
기능성분 함량 및 항산화활성 분석
통계분석
결과 및 고찰
귀리 가수분해물의 일반성분 변화
귀리 가수분해물의 구성 아미노산 조성 및 단백질 품질 특성
귀리 가수분해물의 아베난쓰라마이드 함량 변화
귀리 가수분해물의 총 폴리페놀, 총 플라보노이드 및 항산화 활성 변화
적 요
서 론
최근 한국 사회는 급격한 고령화에 따른 인구 구조의 재편을 경험하고 있으며, 2023년 기준 고령 인구 비중이 18.4%에 육박하여 초고령사회 진입을 목전에 두고 있다(Statistics Korea, 2023). 이와 같은 급격한 고령화는 근감소증(sarcopenia)을 포함한 노인성 질환의 급증을 초래하는 주요 기제로 작용한다. 근감소증은 신체 기능의 퇴행적 변화를 넘어 낙상, 골절 및 만성 대사 질환의 위험을 심화시켜 노년기 삶의 질 저하 및 사회적 의료비용 증대의 주요 요인으로 작용한다(Cruz-Jentoft et al., 2019). 따라서 근감소증의 효과적인 예방 및 관리를 위한 식이 중재 전략의 수립과 함께, 생체 이용률이 극대화된 고기능성 단백질 소재의 개발 및 특성 구명에 관한 연구가 요구된다(Duque-Estrada et al., 2026).
고령층을 위한 효율적인 단백질 공급원의 확보가 요구됨에 따라, 최근 귀리(Avena sativa L.)의 영양학적 가치와 기능적 특성이 주목받고 있다. 귀리는 약 12~20%의 단백질을 함유하고 필수 아미노산 조성이 우수할 뿐만 아니라, 항산화 및 항염증 활성을 지닌 특이적 폴리페놀인 아베난쓰라마이드(avenanthramides, AVN)를 포함하고 있어 노인성 질환 예방을 위한 기능성 소재로서 높은 잠재력을 지닌다(Alemayehu et al., 2023; Xie et al., 2024). 그러나 귀리의 단백질과 유효 성분들은 β-glucan 중심의 복합성 다당류 구조 내에 물리적으로 포집되어 있어, 소화 효소의 접근성 제한에 따른 낮은 생체 이용률이 주요 한계로 지적된다(Alemayehu et al., 2023; Duque-Estrada et al., 2026). 이를 극복하기 위해 열처리 및 압출 성형 등 다양한 가공 기법이 검토되고 있으며, 특히 효소 가수분해는 단백질의 저분자화를 통한 흡수율 향상과 페놀화합물의 유리화를 유도하여 소재의 생리활성 이용 효율을 극대화할 수 있는 유망한 공정으로 보고되고 있다(Nwachukwu & Aluko, 2019; Samtiya et al., 2021).
기존의 귀리 관련 연구는 주로 효소 처리에 따른 귀리의 총 폴리페놀 함량의 증대 및 항산화 활성 변화 등 특정 기능성 성분의 개선에 국한되어 수행되어 왔다(Bei et al., 2018; Streimikyte et al., 2022). 반면, 귀리 단백질에 관한 연구는 유화성, 수분결합력 및 겔 형성능과 같은 가공 적성 및 식품 시스템으로의 적용성 구명에 치중되어 있으며(Klose & Arendt, 2012; Kumar et al., 2021), 단백질의 영양학적 품질과 생체 이용률을 기능성 성분의 변화와 유기적으로 연계하여 평가한 사례는 매우 제한적이다. 특히, 귀리 매트릭스 내에서 단백질의 저분자화와 결합형 페놀화합물의 유리화가 동시에 일어남에도 불구하고, 이들이 소화 과정에서 나타내는 상호작용 및 통합적인 품질 변화에 관한 체계적인 분석은 여전히 미흡한 실정이다. 따라서 고령층의 영양 및 생리적 특성을 고려할 때, 단백질 품질 개선과 기능성 성분의 이용 효율 증대를 동시에 달성하기 위한 통합적 접근이 요구된다.
따라서 본 연구에서는 국내 육성 품종인 ‘대양’ 귀리를 대상으로 효소 가수분해 처리에 따른 일반성분, 기능성 성분 및 단백질 품질 변화를 종합적으로 분석하였다. 이를 통해 가공 공정이 귀리의 품질 특성에 미치는 영향을 구명하고, 최종적으로 귀리를 고령층 맞춤형 고기능성 식물성 단백질 소재로 응용하기 위한 학술적 근거와 전략적 활용 가능성을 제시하고자 하였다.
재료 및 방법
실험 재료
본 연구에서는 2023년 국내에서 수확한 ‘대양’ 품종 귀리를 실험 재료로 사용하였다. 귀리 시료는 정읍명품귀리사업단(Jeongeup, Korea)으로부터 분양받아 사용하였으며, 이물질을 제거한 후 실험 전까지 4°C에서 냉장 보관하였다. 별도의 처리를 거치지 않은 귀리(untreated oat)는 대조군으로서 Control로 명명하였다.
귀리 가수분해물 및 분말 시료 제조
귀리 가수분해물의 조제는 삼육네이처세븐(Sahmyook Nature Seven, Bonghwa, Korea)에 의뢰하여 수행하였다. 분쇄된 귀리 시료는 증숙 및 액화 과정을 통해 전분을 호화시킨 후, 단백질 및 다당류 분해를 위해 복합효소제를 첨가하여 일정 조건에서 가수분해를 진행하였다. 효소 반응 종료 후에는 고온 열처리를 통해 효소를 불활성화하였다. 가수분해 공정을 거친 시료는 후속 처리 방식에 따라 세 가지 형태로 조제되었다. 먼저, 가수분해액을 농축하여 수분을 제거한 농축물(concentrated oat hydrolysate)을 COH로 정의하였다. 분말 형태의 시료는 분무건조 공정을 통해 제조하였으며, 이때 가수분해물 내 불용성 고형분의 제거 여부에 따라 구분하였다. 즉, 가수분해물을 80 mesh 표준망체로 여과하여 청징화한 후 분무건조한 시료(clarified hydrolysate powder)를 CHP로, 여과 공정 없이 전체 가수분해물을 그대로 분무건조한 시료(non-clarified hydrolysate powder)를 NCHP로 각각 명명하였다.
일반성분 분석
수분, 조단백질, 조지방 및 회분 함량은 AOAC 방법(AOAC, 2019)에 준하여 분석하였다. 수분 함량은 시료 1 g을 수분 캔에 취하여 105°C 건조 오븐에서 항량이 될 때까지 건조한 후 측정하였다. 건조 후 시료는 데시케이터에서 방냉한 뒤 무게를 측정하여 수분 함량을 계산하였다.
조단백질 함량은 Micro-Kjeldahl법에 따라 분석하였다. 시료 0.5 g에 황산 10 mL과 촉매제를 첨가하여 분해기(Tecator Digestor auto, Foss, Hillerød, Denmark)를 이용해 420°C에서 1시간 분해하였다. 이후 단백질 분석기(Vapodest 50s, C. Gerhardt GmbH & Co. KG, Königswinter, Germany)를 이용하여 질소 함량을 측정하고, 질소-단백질 환산계수(N×5.83)를 적용하여 조단백질 함량을 산출하였다.
조지방 함량은 Soxhlet 추출법을 이용하여 Soxtherm automatic system (Soxtherm sox416, C. Gerhardt GmbH & Co. KG)으로 분석하였다. 시료 2 g을 n-hexane 용매를 이용하여 추출한 후, 용매를 제거하고 잔류 지방의 무게를 측정하여 함량을 산출하였다.
회분 함량은 시료 1 g을 회화로에서 600°C로 5시간 회화시킨 후 측정하였다. 회화 후 시료는 데시케이터에서 방냉한 뒤 무게를 측정하여 회분 함량을 계산하였다.
탄수화물 함량은 수분, 조단백질, 조지방 및 회분 함량을 합한 값을 100에서 차감하여 계산하였다. 모든 일반성분 결과는 수분 보정을 통해 건물중(dry weight, DW) 기준 g/100 g DW로 환산하여 나타내었다.
구성 아미노산 분석
구성 아미노산 분석은 Han et al.(2026)의 방법에 따라 수행하였다. 트립토판을 제외한 구성 아미노산은 산 가수분해 후 분석하였다. 시료 0.3 g을 밀폐 가능한 유리 시험관에 취하고 6 N HCl 5 mL을 첨가한 후 질소 가스를 주입하여 산소를 제거하였다. 이후 110°C에서 22시간 동안 산 가수분해를 수행하였다. 가수분해 후 시료를 여과하고 증류수를 이용하여 50 mL로 정용하였다. 여과액은 2배 희석한 후 0.22 µm PTFE syringe filter (Merck Millipore, Darmstadt, Germany)로 여과하여 분석 시료로 사용하였다. 아미노산 조성은 아미노산 자동분석기(L-8800, Hitachi High-Tech Co. Tokyo, Japan)를 이용하여 분석하였으며, 이온교환 컬럼(4.6 mm i.d. × 60 mm, 3 µm, Hitachi High-Tech Co.)을 사용하였다. 이동상 조성 및 구배 조건은 제조사에서 제공하는 기본 분석 조건에 따라 수행하였다. 각 아미노산의 정량물질은 표준물질(Amino Acid Mixture Standard Solution Type A, Fujifilm Wako Pure Chemical Co., Osaka, Japan)을 이용하여 분석하였다.
트립토판은 산 가수분해 과정에서 분해되므로 Çevikkalp et al.(2016)의 방법에 따라 별도의 알칼리 가수분해 후 분석하였다. 시료 0.5 g에 5 N sodium hydroxide 20 mL을 첨가하고 질소 가스를 주입한 후, 110°C에서 16시간 동안 가수분해하였다. 가수분해 후 시료를 냉각하여 원심분리(10,000 rpm, 10분)한 뒤 상층액을 취하고, pH 6.3의 완충용액으로 정용하였다. 이후 0.22 µm PVDF syringe filter (Merck Millipore)로 여과하여 분석 시료로 사용하였다. 트립토판 분석은 UPLC system (Acquity UPLC, Waters, Milford, MA, USA)과 photodiode array detector를 이용하여 수행하였다. 컬럼은 Lichrospher 60 RP-Select B (250 mm × 4 mm, 5 µm, Agilent, Santa Clara, CA, USA)를 사용하였으며, 이동상은 ammonium acetate buffer (pH 6.3)와 acetonitrile 혼합용액을 사용하였다. 유속은 1.0 mL/min, 주입량은 10 µL, 검출파장은 280 nm로 설정하였다.
단백질 품질 분석
단백질 품질 평가는 in vitro 단백질 소화율 및 아미노산 조성을 기반으로 단백질 소화율 보정 아미노산가(protein digestibility corrected amino acid score, PDCAAS)를 산출하여 평가하였다(Schaafsma, 2000). 단백질 소화율은 protein digestibility assay kit (K-PDCAAS, Megazyme, Chicago, IL, USA)를 이용하여 제조사의 방법에 따라 분석하였다. 시료 500 mg에 0.06 N HCl을 첨가하여 37°C에서 전처리한 후, pepsin을 첨가하여 37°C에서 효소반응을 수행하였다. 이후 tris buffer를 이용하여 pH를 중성으로 조정한 뒤 trypsin/chymotrypsin 혼합 효소를 첨가하여 추가 반응을 진행하였다. 효소 반응 후 반응액에 trichloroacetic acid (TCA)를 첨가하여 단백질을 침전시키고, 원심분리(3,000 rpm, 10분)하여 상층액을 회수하였다. 회수된 상층액에 존재하는 유리 아미노산은 ninhydrin 반응을 통해 정량하였으며, 분광광도계(EPOCH2, Bio Tek Instruments Inc., Winooski, VT, USA)를 이용하여 520 nm에서 흡광도를 측정하였다. 단백질 소화율은 표준물질 (L-glycine) 및 대조군을 포함하여 계산하였다. 아미노산가(amino acid score)는 구성 아미노산 분석 결과를 바탕으로 산출하였으며, 제1 제한 아미노산(first limiting amino acid)을 기준으로 평가하였다. 최종적으로 단백질 소화율과 아미노산가를 이용하여 protein digestibility-corrected amino acid score (PDCAAS) 값을 산출하였다. 모든 계산은 Megazyme사에서 제공하는 Mega-Calc 프로그램을 이용하여 수행하였다.
아베난쓰라마이드 함량 분석
AVN 함량 분석을 위해 시료 1 g에 80% ethanol (0.01 M phosphate buffer, pH 2.8) 10 mL을 첨가하여 37°C에서 240 rpm으로 16시간 추출하였다. 여과 후 동일 용매를 이용하여 37°C에서 20분 동안 재추출을 2회 수행하였으며, 회수된 추출물은 감압농축하였다. 농축물은 80% ethanol 2 mL에 재용해 한 후, 0.22 µm PVDF syringe filter (Merck Millipore)로 여과하여 분석 시료로 사용하였다.
AVN은 Acquity UPLC BEH C18 (2.1 × 100 mm, 1.7 µm, Waters) 컬럼을 이용하여 UPLC (Acquity UPLC, Waters)로 정량하였다. 이동상은 0.01 M phosphate buffer (pH 2.8)용액(A)과 acetonitrile (B)을 사용하였다. 용매 농도 구배 조건은 초기 85:15 (A:B, v/v)에서 시작하여 10분 동안 20:80 (A:B)까지 선형적으로 증가시킨 후, 11분에 초기 조건으로 회귀하여 컬럼을 재평형하였다. 유속은 0.6 mL/min, 시료 주입량은 10 µL, 검출 파장은 340 nm로 설정하였다. 각 AVN 성분(AVN-A, AVN-B 및 AVN-C)은 표준물질의 머무름 시간과 비교하여 확인하였으며, 각각의 검량곡선을 이용하여 정량하였다.
기능성분 함량 및 항산화활성 분석
기능성분 함량 및 항산화활성 분석은 Han 등(2026)의 방법에 따라 수행하였다. 시료 3 g에 70% ethanol 30 mL을 가하여 25°C에서 24시간 동안 진탕하여 기능성분을 추출하였다. 추출액은 원심분리(12,000 rpm, 20분, 4°C)하여 침전물을 제거한 후 상층액을 회수하였다. 상층액에 n-hexane 15 mL을 첨가하여 5분간 교반한 후 원심분리(3,000 rpm, 5분, 4°C)하여 상층의 헥산층을 제거하였다. 이 과정을 3회 반복하여 추출물 내 지질 성분을 제거하였다. 이후 ethanol 층을 여과한 뒤 감압농축하여 용매를 제거하고, 잔사를 회수하여 추출 수율(%)을 산출하였다. 잔사는 dimethyl sulfoxide에 용해(100 mg extract/mL)하여 분석 시료로 사용하였다.
총 폴리페놀 함량(total polyphenol content, TPC)은 Folin-Ciocalteu 방법에 따라 측정하였다. 추출물 10 µL에 2% Na2CO3 200 µL를 가하여 3분 동안 반응시킨 후, 50% Folin-Ciocalteu 시약 10 µL 첨가하여 27분 동안 암조건에서 반응을 진행하였다. 이후 microplate reader (Elx 808, BioTek Instruments Inc., Winooski, VT, USA)를 이용하여 750 nm에서 반응액의 흡광도를 측정하였으며, gallic acid를 표준물질로 하여 검량곡선을 작성한 후 mg gallic acid equivalents (mg GAE/100 g DW)로 나타내었다. 총 플라보노이드 함량(total flavonoid content, TFC)은 aluminum chloride colorimetric 방법에 따라 분석하였다. 추출물 25 µL에 증류수 100 µL와 5% NaNO2 7.5 µL를 가하여 5분 동안 반응시킨 후, 10% AlCl3·6H2O 15 µL를 첨가하여 6분 동안 암조건에서 반응을 진행하였다. 이후 반응액에 1 N NaOH 50 µL을 가하여 11분 동안 반응시킨 후, microplate reader (Elx 808, BioTek Instruments Inc.)를 이용하여 510 nm에서 반응액의 흡광도를 측정하였다. Catechin을 표준물질로 하여 검량곡선을 작성한 후 mg catechin equivalents (mg CE/100 g DW)로 나타내었다.
항산화활성은 2,2-diphenyl-1-picrylhydrazyl (DPPH) 및 2,2’-azino-bis(3-tehylbenzothiazoline-6-sulfonic acid) (ABTS) 라디칼 소거능과 ferric reducing antioxidant power (FRAP)를 평가하였다. DPPH 및 ABTS 라디칼 소거능은 추출물 20 µL에 각각 0.2 mM DPPH 및 7.4 mM ABTS 용액 200 µL를 첨가하여 30분 동안 반응시킨 후, microplate reader (Elx 808, BioTek Instruments Inc.)를 이용하여 각각 517 nm 및 734 nm에서 반응액의 흡광도를 측정하였다. DPPH 및 ABTS 소거능은 Trolox를 표준물질로 하여 검량곡선을 작성한 후 mg Trolox equivalents (mg TE/g DW)로 나타내었다. FRAP은 추출물 6 µL에 FRAP reagent 180 µL와 증류수 18 µL를 혼합한 후, 10분 동안 암조건에서 반응을 진행시켰다. 이후 microplate reader (Elx 808, BioTek Instruments Inc.)를 이용하여 593 nm에서 반응액의 흡광도를 측정하였으며, mM Fe2+/g DW로 나타내었다.
통계분석
모든 실험은 3회 반복 수행하였으며, 결과는 평균과 표준편차로 나타내었다. 통계분석은 SPSS 프로그램(version 18.0, SPSS Inc., Chicago, IL, USA)을 이용하여 수행하였다. 시료 간 유의성은 일원분산분석(one-way analysis of variance, ANOVA)을 실시하였으며, 유의적인 차이가 있는 경우 Duncan의 다중범위검정(Duncan’s multiple range test)을 이용하여 평균 간 차이를 비교하였다. 통계적 유의수준은 p < 0.05로 설정하였다.
결과 및 고찰
귀리 가수분해물의 일반성분 변화
효소 가수분해 및 후속 공정에 따른 귀리 가수분해물의 일반성분 변화는 Table 1에 나타내었다. Control의 조단백질, 조지방, 조회분 및 조탄수화물 함량은 각각 12.03, 12.44, 2.08 및 73.45 g/100 g DW로 분석되었다. 국내 귀리의 일반성분에 대한 선행연구에서 재배 지역 및 연차에 따라 조단백 함량은 9.3~13.7%, 조지방은 8.3~10.6%, 조회분은 1.6~1.8% 및 조탄수화물은 65.7~69.7% 범위로 보고된 바 있다(Song et al., 2023). 본 연구의 Control은 조단백질 함량이 선행연구 범위 내에 있었고, 조지방 및 조탄수화물 함량은 다소 높은 수준을 보였다. 이러한 차이는 품종, 재배 환경, 수확 연도 및 분석 기준의 차이에 기인한 것으로 판단된다.
처리군 간 비교에서 조단백질 함량은 NCHP에서 12.63 g/100 g DW로 가장 높았으며, Control과 유사한 수준을 보였다. 반면 COH와 CHP에서는 각각 6.11 및 3.25 g/100 g DW로 감소하였다. 특히 CHP에서 단백질 함량이 크게 감소한 것은 여과 공정 중 불용성 고형물이 제거되면서 단백질 또는 단백질 함유 입자가 함께 손실되었기 때문으로 판단된다. 반면 NCHP는 여과 공정을 거치지 않고 전체 가수분해물을 분무건조하였기 때문에 단백질 함량이 유지된 것으로 보인다. 조지방 함량은 Control에서 가장 높았고, COH와 NCHP에서는 각각 5.02 및 5.31 g/100 g DW 수준으로 감소하였으며, CHP에서는 0.09 g/100 g DW로 가장 낮았다. 조탄수화물 함량은 CHP에서는 94.72 g/100 g DW로 가장 높았는데, 이는 단백질 및 지질 함량 감소에 따른 상대적 증가에 기인한 것으로 해석된다. 이러한 결과는 효소 가수분해 및 분획 공정이 귀리의 일반성분 조성에 유의적인 영향을 미치며, 특히 고형분 제거 여부가 단백질 및 지질 함량 변화에 중요한 요인으로 작용함을 시사한다.
Table 1.
Proximate composition of oat samples subjected to enzymatic hydrolysis and processing (g/100 g dry weight).
| Treatments | Protein | Lipid | Ash | Carbohydrate |
| Control1) | 12.03 ± 0.05b2) | 12.44 ± 0.23a | 2.08 ± 0.03a | 73.45 ± 0.22d |
| COH | 6.11 ± 0.05c | 5.02 ± 0.05c | 1.35 ± 0.05d | 87.51 ± 0.09b |
| CHP | 3.25 ± 0.01d | 0.09 ± 0.01d | 1.94 ± 0.01b | 94.72 ± 0.02a |
| NCHP | 12.63 ± 0.35a | 5.31 ± 0.04b | 1.86 ± 0.01c | 80.20 ± 0.38c |
효소 가수분해 공정은 효소 농도 및 반응 조건에 따라 곡류의 일반성분 조성에 영향을 미칠 수 있는 것으로 보고되고 있다. 유사한 곡류 기반 효소 처리 연구에서도 단백질 및 지질 함량이 변화하는 것으로 보고된 바 있으며(Aenglong et al., 2024), 본 연구에서도 효소 가수분해 이후의 여과 및 건조 공정에 따라 일반성분 조성이 상이하게 나타났다. 한편 효소 가수분해는 일반성분 변화뿐 아니라 곡류 매트릭스의 용해성, 점도 및 분산성에도 영향을 미치는 것으로 알려져 있으며, 귀리겨를 효소 가수분해한 연구에서는 영양적 품질을 유지하면서 음료 적용 시 용해성 및 물성 개선 효과가 확인된 바 있다(Tan et al., 2023).이러한 결과는 효소 가수분해 및 후속 공정이 귀리의 영양학적 특성 변화에 영향을 미칠 수 있음을 보여주며, 향후 용해성, 분산성 및 물성과 같은 가공적성에 대한 추가적인 연구가 필요할 것으로 판단된다.
귀리 가수분해물의 구성 아미노산 조성 및 단백질 품질 특성
귀리 가수분해물의 구성 아미노산 조성은 Table 2에 나타내었다. 일반적으로 귀리 단백질은 glutamic acid, aspartic acid 및 leucine 함량이 상대적으로 높은 것으로 알려져 있으며, 곡류 중에서는 비교적 우수한 필수 아미노산 조성을 갖는 것으로 보고되고 있다(Klose & Arendt, 2012). 본 연구에서도 모든 처리구에서 glutamic acid 함량이 가장 높았으며, aspartic acid, leucine 및 serine이 비교적 높은 수준을 보였다. 반면, methionine 및 tryptophan 함량은 상대적으로 낮았다. 총 아미노산 함량은 Control에서 12.41 g/100 g DW로 가장 높았고, NCHP, COH 및 CHP 순이었다. 특히 CHP에서는 대부분의 구성 아미노산 함량이 크게 감소하였는데, 이는 여과 공정 중 단백질 및 질소화합물을 포함한 불용성 고형분이 제거되었기 때문으로 판단된다. 반면, NCHP는 여과 공정을 거치지 않았기 때문에 glutamic acid, leucine 및 proline 등의 함량이 Control 수준으로 유지되었다. 필수 아미노산 비율은 Control과 COH에서 각각 0.33 수준으로 가장 높았으며, NCHP는 0.31, CHP는 0.25였다. BCAA 함량 또한 Control에서 1.82 g/100 g DW로 가장 높았고, NCHP, COH 및 CHP 순으로 감소하였다. 특히 leucine, isoleucine 및 valine으로 구성되는 BCAA는 근육 단백질 합성과 관련된 주요 아미노산으로 알려져 있어(Devries and Phillips, 2015), NCHP는 상대적으로 높은 BCAA 함량을 유지하였다. 한편, sulfur-containing amino acid (SAA) 함량은 Control에서 가장 높았으며, 처리구에서는 전반적으로 감소하는 경향을 보였다. Lysine 함량은 모든 처리구에서 상대적으로 낮게 유지되었으며, 이는 이후 단백질 품질 평가에서 lysine이 제1 제한 아미노산으로 확인된 결과와도 일치하였다(Table 3).
Table 2.
Constituent amino acid content (g/100 g dry weight) of oat sample subjected to enzymatic hydrolysis and processing.
| Amino acids | Control1) | COH | CHP | NCHP |
| Alanine | 0.66 ± 0.01a2) | 0.42 ± 0.01c | 0.13 ± 0.00d | 0.53 ± 0.01b |
| Arginine | 0.77 ± 0.01a | 0.46 ± 0.01c | 0.07 ± 0.00d | 0.56 ± 0.01b |
| Aspartic acid | 1.13 ± 0.02a | 0.71 ± 0.01c | 0.27 ± 0.00d | 0.96 ± 0.01b |
| Cysteine | 0.47 ± 0.00a | 0.33 ± 0.00c | 0.23 ± 0.00d | 0.36 ± 0.00b |
| Glutamic acid | 3.00 ± 0.03a | 1.93 ± 0.03c | 0.67 ± 0.00d | 2.95 ± 0.02b |
| Glycine | 0.66 ± 0.01a | 0.43 ± 0.01c | 0.18 ± 0.00d | 0.56 ± 0.00b |
| Histidine | 0.29 ± 0.01a | 0.18 ± 0.00c | 0.03 ± 0.00d | 0.23 ± 0.01b |
| Isoleucine | 0.35 ± 0.01a | 0.22 ± 0.01c | 0.04 ± 0.00d | 0.29 ± 0.01b |
| Leucine | 0.92 ± 0.01a | 0.57 ± 0.02c | 0.06 ± 0.00d | 0.79 ± 0.01b |
| Lysine | 0.53 ± 0.01a | 0.28 ± 0.01c | 0.08 ± 0.00d | 0.33 ± 0.00b |
| Methionine | 0.12 ± 0.00a | 0.07 ± 0.02b | 0.00 ± 0.01c | 0.05 ± 0.01b |
| Phenylalanine | 0.68 ± 0.01a | 0.44 ± 0.01c | 0.14 ± 0.01d | 0.60 ± 0.00b |
| Proline | 0.61 ± 0.03a | 0.34 ± 0.01c | 0.08 ± 0.00d | 0.65 ± 0.01b |
| Serine | 0.76 ± 0.01a | 0.50 ± 0.01c | 0.23 ± 0.00d | 0.67 ± 0.00b |
| Threonine | 0.48 ± 0.01a | 0.31 ± 0.01c | 0.12 ± 0.00d | 0.40 ± 0.00b |
| Tryptophan | 0.16 ± 0.00b | 0.14 ± 0.00c | 0.07 ± 0.00d | 0.16 ± 0.00a |
| Tyrosine | 0.28 ± 0.00a | 0.17 ± 0.01c | 0.10 ± 0.00d | 0.22 ± 0.00b |
| Valine | 0.55 ± 0.01a | 0.37 ± 0.02c | 0.09 ± 0.00d | 0.45 ± 0.02b |
| TAA3) | 12.41 ± 0.17a | 7.88 ± 0.20c | 2.58 ± 0.01d | 10.74 ± 0.06b |
| EAA/TAA4) | 0.33 ± 0.00a | 0.33 ± 0.01a | 0.25 ± 0.00c | 0.31 ± 0.00b |
| BCAA5) | 1.82 ± 0.04a | 1.16 ± 0.06c | 0.19 ± 0.00d | 1.52 ± 0.03b |
| SAA6) | 0.59 ± 0.00a | 0.40 ± 0.02b | 0.23 ± 0.01c | 0.41 ± 0.01b |
1)Control, untreated oat; COH, concentrated oat hydrolysate; CHP, clarified hydrolysate powder; NCHP, non-clarified hydrolysate powder.
2)Values are expressed as the mean ± standard deviation (n = 3). a-dMeans with different letters within the same column are significantly different (p < 0.05) according to Duncan’s multiple range test.
구성 아미노산 조성 변화와 함께 귀리 가수분해물의 실제 단백질 이용성을 평가하기 위하여 in vitro digestibility, amino acid score 및 PDCAAS를 분석하였다(Table 3). In vitro digestibility는 단백질의 소화가능성을 나타내는 지표로, 값이 높을수록 효소에 의해 단백질이 보다 쉽게 분해됨을 의미한다(Krul et al., 2024). 일반적으로 곡류 및 식물성 단백질의 in vitro digestibility는 원료의 조성 및 가공 조건에 따라 달라질 수 있으며, 열처리 및 분획 공정은 단백질의 구조적 접근성과 효소 반응성에 영향을 미치는 것으로 알려져 있다. 또한 효소 가수분해 이후의 농축 및 분무건조 과정은 단백질의 재응집 또는 구조적 변화를 유도하여 소화율에 영향을 미칠 수 있는 것으로 보고되고 있다(Fu et al., 2023; Santos-Sánchez et al., 2024). 본 연구에서는 in vitro digestibility는 Control에서 1.01로 가장 높았으며, 처리구에서는 0.84~0.88 수준을 보여 다소 감소하는 경향을 보였다. 그러나 모든 처리구가 0.84 이상의 값을 유지하여 전반적으로 양호한 소화 특성을 나타내는 것으로 판단된다. 모든 처리구에서 lysine이 제1 제한 아미노산으로 확인되었으며, 이는 구성 아미노산 분석에서 lysine 함량이 상대적으로 낮았던 결과와 일치하였다. Amino acid score는 NCHP에서 가장 높았고, CHP, Control 및 COH 순이었다. PDCAAS는 단백질의 아미노산 조성과 소화율을 동시에 반영하는 대표적인 단백질 품질 평가 지표로, 값이 높을수록 인체 내 이용성이 우수한 단백질임을 의미한다(Schaafsma, 2000). 일반적으로 식물성 단백질의 PDCAAS는 원료 특성 및 가공 조건에 따라 다양한 범위를 보이며, 곡류 단백질은 lysine 제한성으로 인해 상대적으로 낮은 값을 나타내는 것으로 알려져 있다(Joye, 2019; Sá & House, 2024). 본 연구에서는 PDCAAS가 NCHP에서 0.42로 가장 높았으며, CHP (0.33), Control (0.26) 및 COH (0.17) 순이었다. 이는 NCHP가 필수 아미노산 조성과 소화율을 함께 고려한 단백질 품질 측면에서 가장 우수한 특성을 보였음을 의미한다.
Table 3.
In vitro protein digestibility, limiting amino acid, amino acid score, and PDCAAS of oat samples subjected to enzymatic hydrolysis and processing.
| Treatments | In vitro digestibility | First limiting amino acid | Amino acid score | PDCAAS |
| Control1) | 1.01 | L-Lysine | 0.26 | 0.26 |
| COH | 0.84 | L-Lysine | 0.20 | 0.17 |
| CHP | 0.86 | L-Lysine | 0.39 | 0.33 |
| NCHP | 0.88 | L-Lysine | 0.48 | 0.42 |
구성 아미노산 조성과 단백질 품질 평가 결과를 종합하면, 효소 가수분해 및 후속 공정에 따라 귀리 단백질의 영양학적 특성이 서로 다른 양상을 보였다. 특히 NCHP는 구성 아미노산 함량을 비교적 잘 유지하였고, amino acid score와 PDCAAS에서도 가장 높은 값을 보여 실질적인 단백질 품질 측면에서 가장 우수한 특성을 보였다. 반면 CHP는 여과 공정에 의해 단백질 및 아미노산 함량이 크게 감소하였으며, COH는 농축 공정 이후 단백질 품질 지표가 상대적으로 낮았다. 이러한 결과는 귀리 단백질의 품질 평가에서 단순 소화율뿐 아니라 필수 아미노산 조성과 소화율을 함께 고려한 종합적인 평가가 필요함을 보여준다.
귀리 가수분해물의 아베난쓰라마이드 함량 변화
AVN은 귀리에 특이적으로 존재하는 phenolic alkaloid계 화합물로, 항산화 및 항염 활성이 우수한 기능성 성분으로 알려져 있다. 일반적으로 귀리에는 AVN-A, AVN-B 및 AVN-C가 주요 형태로 존재하며, 품종, 재배 환경 및 가공 조건에 따라 함량 차이가 발생하는 것으로 보고되었다(Xie et al., 2024). 또한 귀리의 총 AVN 함량은 약 25~407 µg/g DW 범위로 보고되었으며, AVN-B와 AVN-C가 상대적으로 높은 비율을 차지하는 경향이 확인된 바 있다(Dvořáček et al., 2021). 본 연구에서 귀리 가수분해물의 AVN 함량 변화를 Table 4에 나타내었다. Control의 총 AVN 함량은 152.75 µg/g DW였으며, AVN-B, AVN-C 및 AVN-A 순으로 높은 함량을 보였다. 반면 NCHP에서는 총 AVN 함량이 308.79 µg/g DW로 가장 높아 Control 대비 약 2.02배 증가하였다. 특히 AVN-A와 AVN-B의 증가폭이 크게 나타났으며, AVN-C 또한 유의적으로 증가하였다(p < 0.05). 반면 CHP에서는 총 AVN 함량이 74.43 µg/g DW로 감소하였고, COH에서는 56.40 µg/g DW로 가장 낮은 값을 보였다. 이러한 결과는 효소 가수분해 과정에서 결합형 AVN의 유리화가 촉진된 반면, 여과 공정에서는 일부 AVN이 불용성 고형분과 함께 제거되었기 때문으로 판단된다. 또한 농축 공정에서는 열 및 공정 조건에 의해 AVN의 손실 또는 분해가 발생했을 가능성이 있는 것으로 사료된다.
Table 4.
Avenanthramides (AVN) content of oat samples subjected to enzymatic hydrolysis and processing (μg/g dry weight).
| Treatments | AVN-A | AVN-B | AVN-C | Total |
| Control1) | 36.30 ± 0.17b2) | 63.05 ± 1.54b | 53.40 ± 0.08b | 152.75 ± 1.69b |
| COH | 22.12 ± 0.16d | 24.31 ± 0.09d | 9.97 ± 0.13d | 56.40 ± 0.26d |
| CHP | 24.35 ± 0.19c | 31.82 ± 0.51c | 18.25 ± 0.09c | 74.43 ± 0.34c |
| NCHP | 104.47 ± 1.15a | 132.38 ± 0.70a | 71.95 ± 0.87a | 308.79 ± 2.65a |
귀리 가수분해물의 총 폴리페놀, 총 플라보노이드 및 항산화 활성 변화
귀리 가수분해물의 TPC 및 TFC는 Fig. 1에 나타내었다. 일반적으로 귀리의 페놀화합물은 세포벽 다당류 및 단백질과 결합된 형태로 존재하며, 효소 가수분해를 통해 결합형 페놀화합물의 유리화가 촉진될 수 있는 것으로 보고되고 있다(Bei et al., 2018; Streimikyte et al., 2022). 본 연구에서도 효소 처리군은 Control 대비 TPC 및 TFC가 전반적으로 증가하는 경향을 보였다.
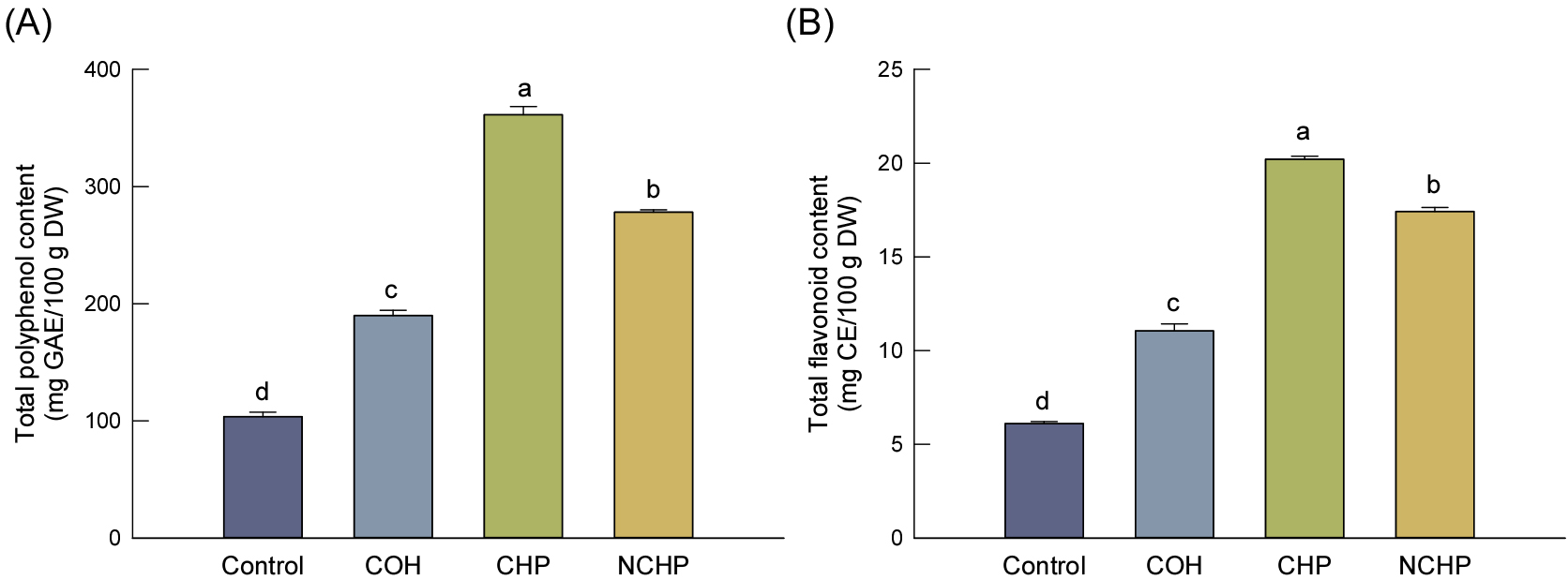
Fig. 1
Total polyphenol (A) and flavonoid (B) contents of oat samples subjected to enzymatic hydrolysis and processing. Total polyphenol and flavonoid contents are expressed as milligram gallic acid equivalents (GAE) and milligram catechin equivalents (CE), respectively, per 100 gram of dry weight (DW). Values are expressed as the mean ± standard deviation (n = 3). Different letters above the bars indicate significant differences (p < 0.05) according to Duncan’s multiple range test. Control, untreated oat; COH, concentrated oat hydrolysate; CHP, clarified hydrolysate powder; NCHP, non-clarified hydrolysate powder.
TPC는 CHP에서 361.30 mg GAE/100 g DW로 가장 높았으며, 이는 Control (103.58 mg GAE/100 g DW) 대비 약 3.49배 증가한 수준이었다. TFC 또한 CHP에서 20.21 mg CE/100 g DW로 가장 높았고, NCHP, COH 및 Control 순으로 감소하였다. 특히 COH보다 CHP와 NCHP에서 높은 함량을 나타낸 것은 농축 공정보다 분무건조 공정이 기능성 성분 유지에 보다 유리하였기 때문으로 판단된다. 반면 여과 여부에 따라서는 CHP가 NCHP보다 높은 값을 보였는데, 이는 여과 공정 중 불용성 고형분 및 추출 간섭 물질이 제거됨에 따라 페놀화합물의 추출 효율 및 검출 특성이 향상되었기 때문으로 사료된다. 한편, AVN 함량은 NCHP에서 가장 높아(Table 4) TPC 및 TFC와는 상이한 경향을 보였는데, 이는 특정 기능성 성분과 총 페놀화합물 간의 공정 민감성 차이에 기인한 것으로 판단된다. TPC 및 TFC 분석은 다양한 페놀성 화합물과 환원성 물질의 총량을 반영하는 반면, AVN은 귀리에 특이적으로 존재하는 개별 phenolic amide 화합물을 정량한 결과이기 때문에 공정에 따른 변화 양상이 다르게 나타날 수 있다. 특히 여과 공정은 불용성 고형분 제거를 통해 총 페놀성 화합물의 추출 효율 및 분석 특성을 향상시킬 수 있으나, 일부 AVN은 고형분과 함께 제거되어 감소한 것으로 판단된다.
귀리 가수분해물의 항산화 활성은 Fig. 2에 나타내었다. DPPH 및 ABTS 라디칼 소거능과 FRAP 모두 CHP에서 가장 높았으며, 각각 0.93 mg TE/g DW, 4.46 mg TE/g DW 및 7.97 mM/g DW 수준을 보였다. 반면 Control에서는 각각 0.25 mg TE/g DW, 1.16 mg TE/g DW 및 0.77 mM/g DW로 가장 낮았다. 이러한 경향은 TPC 및 TFC 변화와 유사하였으며, 귀리 가수분해물의 항산화 활성이 페놀화합물 함량과 밀접한 관련이 있음을 보여준다. 선행연구에서도 귀리 유래 페놀화합물 및 AVN이 우수한 항산화 활성을 보이며, 효소 처리에 의해 항산화 활성이 증가할 수 있음이 보고된 바 있다(Xie et al., 2024).
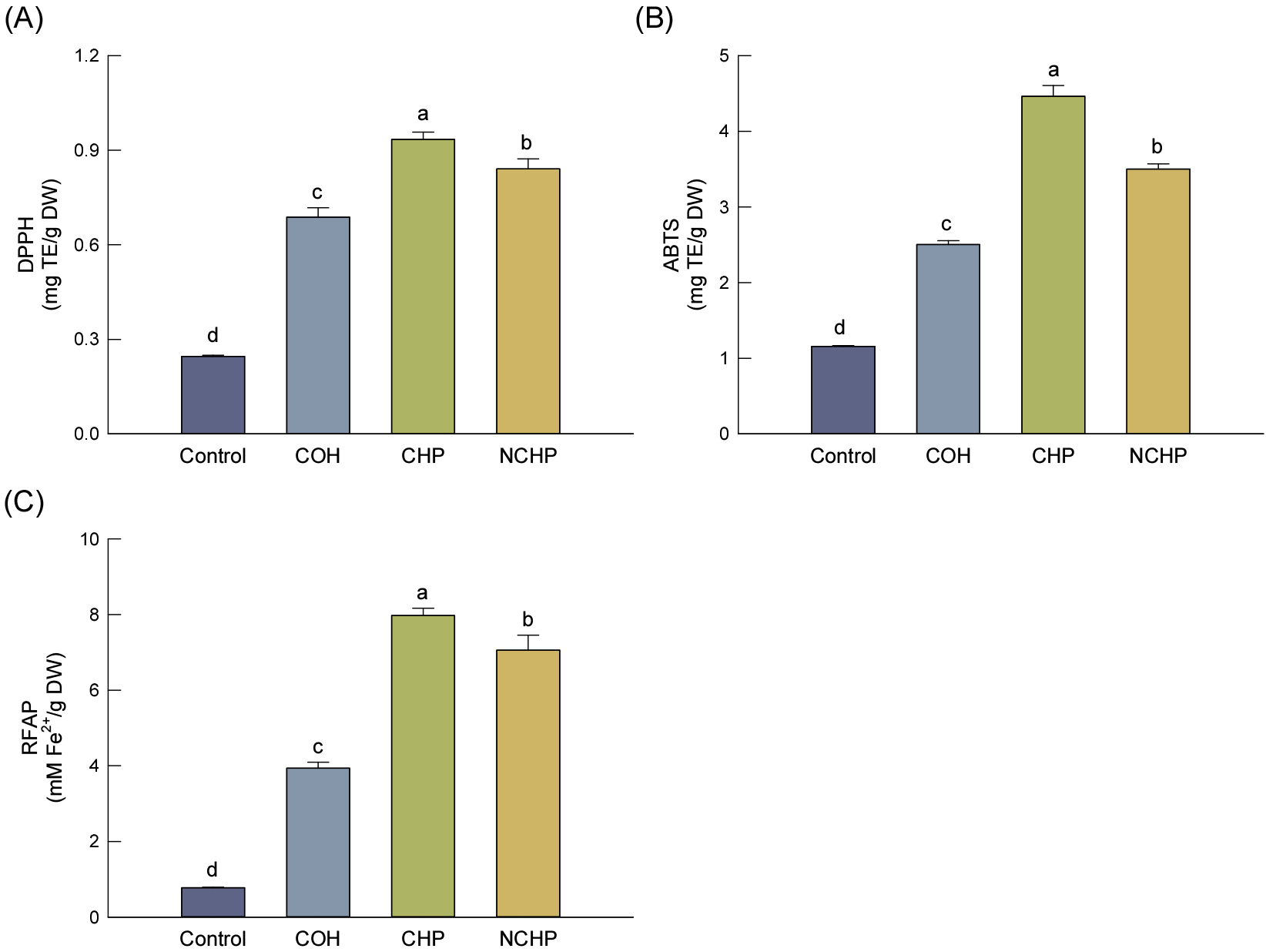
Fig. 2
Antioxidant activities of oat samples subjected to enzymatic hydrolysis and processing, including radical scavenging activities of 2,2-diphenyl-1-picrylhydrazyl (DPPH) (A), 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) (B), and ferric reducing antioxidant power (FRAP) (C). DPPH and ABTS activities are expressed as milligram Trolox equivalents (TE) per gram of dry weight (DW), whereas FRAP is expressed as mM Fe2+/g DW. Values are expressed as the mean ± standard deviation (n = 3). Different letters above the bars indicate significant differences (p < 0.05) according to Duncan’s multiple range test. Control, untreated oat; COH, concentrated oat hydrolysate; CHP, clarified hydrolysate powder; NCHP, non-clarified hydrolysate powder.
종합적으로, 효소 가수분해 및 후속 공정은 귀리의 단백질 품질과 기능성 성분 조성에서 서로 다른 영향을 미치는 것으로 나타났다. 특히 후속 공정 중 여과 공정은 단백질, 구성 아미노산 및 AVN 함량 감소에 가장 큰 영향을 미친 것으로 판단된다. 실제로 여과 공정을 거친 CHP에서는 조단백질, 총 아미노산 및 AVN 함량이 감소한 반면, 비여과 처리구인 NCHP에서는 상대적으로 높은 수준이 유지되었다. TPC 및 TFC는 CHP에서 가장 높게 나타났는데, 이는 여과 과정에서 불용성 고형분 및 추출 간섭 물질이 제거되면서 총 페놀화합물의 추출 효율 및 검출 특성이 향상되었기 때문으로 해석된다. NCHP는 상대적으로 높은 PDCAAS 및 AVN 함량을 보여 단백질 품질과 귀리 특이 기능성 성분 측면에서 우수한 특성을 보였다. CHP는 높은 TPC 및 TFC와 항산화 활성을 보여 항산화 기능성 소재로서의 활용 가능성을 확인하였다. 이러한 결과는 귀리 가수분해물의 기능성 특성이 효소 처리 자체뿐 아니라 후속 분획 및 건조 공정에 의해 크게 영향을 받을 수 있음을 시사하며, 목적에 따라 단백질 품질 중심 또는 기능성 중심의 맞춤형 식물성 소재 개발이 가능함을 보여준다.
적 요
본 연구에서는 국내산 ‘대양’ 귀리를 대상으로 효소 가수분해 및 후속 공정에 따른 단백질 품질과 기능성 특성을 평가하여 식물성 단백질 소재로서의 활용 가능성을 검토하고자 하였다. 시료는 원곡(untreated oat, Control), 농축 가수분해물(concentrated oat hydrolysate, COH), 여과 후 분무건조 분말(clarified hydrolysate powder, CHP), 비여과 분무건조 분말(non-clarified hydrolysate powder, NCHP)로 구분하여 분석하였다. 조단백질 함량은 COH와 CHP에서 감소한 반면, NCHP에서는 Control과 유사한 수준을 유지하였다. 총 아미노산 함량과 BCAA 함량 또한 NCHP에서 상대적으로 높게 유지되었으며, 모든 처리구에서 lysine이 제1 제한 아미노산으로 확인되었다. In vitro digestibility는 처리구에서 다소 감소하였으나 전반적으로 양호한 수준을 유지하였으며, PDCAAS는 NCHP에서 0.42로 가장 높았다. 총 아베난쓰라마이드(avenanthramides, AVN) 함량은 NCHP에서 308.79 µg/g DW로 가장 높았으며, COH에서 가장 낮았다. 반면 총 폴리페놀 및 플라보노이드 함량과 항산화 활성은 CHP에서 가장 우수하였다. 이상의 결과를 종합하면, 효소 가수분해 및 후속 공정은 귀리의 단백질 품질과 기능성 특성에 서로 다른 영향을 미치는 것으로 확인되었다. 특히 NCHP는 단백질 품질 및 귀리 특이 기능성 성분 측면에서 우수한 특성을 보였으며, CHP는 항산화 기능성 소재로서의 활용 가능성을 보였다. 이러한 결과는 후속 공정 조건에 따라 목적 지향적인 귀리 기반 식물성 소재 개발이 가능함을 시사한다.
