

이화명나방(Chilo suppressalis Walker, Rice stem borer) 은 유충이 벼를 중심으로 피, 갈대, 줄풀 등 벼과 식물을 가 해하는 해충으로서 우리나라뿐만 아니라 동남아시아, 중국, 일본, 및 인도까지 광범위하게 분포하는 대표적인 해충 중 의 하나이다. 이화명나방은 일반적으로 추운 북부지방을 제 외한 우리나라 대부분의 지방에서는 연 2회 발생하며, 5월 경에 1세대 성충이 우화하고, 2세대는 7월경에 우화한다. 그 이후 유충은 볏짚이나 벼 그루터기에서 월동한다고 알려져 있다(Atapour & Moharramipour, 2009; Lee & Park, 1991). 특히 우리나라에서는 1960년대 후반까지 전국적으로 논에 서 매년 대발생하여 그 피해가 심각하였다. 그러나 1970년 대 중반 이후 벼의 조기 이앙, 조기 수확, 다비·밀식, 집중약 제방제 및 기계수확 등의 요인으로 인하여 이화명나방 유충 의 발육부진 등의 결과로 이어졌고 유충의 월동율이 낮아지 거나 사망률이 높아져 발생량이 격감하였고, 그 이후 국내 방제 대상해충에서 제외되었다(Lee & Park, 1991; Lee 1999).
억새(Miscanthus)는 화본과에 속하는 C4 식물로서 한국, 중국, 일본 및 동남아시아에 분포하는 대표적인 섬유질계 바이오에너지작물 중의 하나이다(An et al., 2013; Atkinson, 2009; Greef et al., 1997; Jezowski, 2008; Koonin, 2006; Lewandowski et al., 2000). 억새는 영년생 식물로서 생산 에 투입되는 에너지 대비 산출되는 에너지가 많으며 질소 이용효율이 높아 무비재배가 가능하며 저온, 건조 등의 열 악환경 및 병해충에 대한 내성이 높다는 장점을 가지고 있 다(Beale & Long, 1995; Lewandowski & Schmidit, 2006; Moon et al., 2010). 최근 농촌진흥청에서 개발한 한국형 바 이오에너지작물 ‘거대 1호’(Miscanthus saccharilforus cv. Geodae 1)는 물억새의 일종으로 생산량이 ha 당 최대 30톤 이나 되어 유럽, 미국 등에서 바이오에탄올 생산용으로 재 배하고 있는 3배체 억새(M. × giganteus)와 비교하여 뒤지 지 않고 습지에서도 생육이 양호하여, 바이오에너지 생산 산업 원료로 국내 토종 억새 유전자원을 이용할 수 있다는 기대감이 커지고 있다(Moon et al., 2010).
최근 거대 1호(M. saccharilforus cv. Geodae 1)를 집단 재배하는 시험포장의 일부에서 억새 줄기를 가해하는 이화 명나방 유충이 발견되었다. 국내외적으로 억새의 병충해에 대한 보고는 거의 없었으나 최근에 Kang et al. (2014)이 거 대억새 대규모 재배 단지 내에 서식하는 곤충상을 조사한 결과, 노린재목 중 멸구류 등이 관찰되었다고 보고하고 있 다. 또한 Jeffrey et al. (2010)에 의하면 바이오에너지작물 인 3배체 억새(M. × giganteus)에서 매미목 진딧물과의 Sipha flava (Forbes)와 Rhopalosiphum maidis (Fitch)을 발견하였 다고 보고하고 있으며 경제적 바이오에너지 작물의 생산성 을 위하여 해충 관리에 대한 논의의 필요성을 시사하였다.
국내에서도 바이오에너지 원료생산을 위해 억새를 대규 모로 재배할 가능성이 높아짐에 따라 억새의 생육 및 수량 감소에 영향을 미치는 잠재적 병충해 발생을 사전에 예방하 며 효과적으로 억제할 수 있는 방법 개발이 절실하다. 본 연 구는 억새 집단 재배지 내의 이화명나방 유충의 월동양상 및 밀도를 조사하여 억새 가해해충 이화명나방의 방제법에 대한 기초자료로 활용하고자 수행하였다.
재료 및 방법
본 연구는 억새에 기주하는 이화명나방의 발생형태 및 밀 도를 분석하여 이화명나방의 방제 및 관리법 개발의 기초로 활용하기 위해 농촌진흥청 국립식량과학원 바이오에너지작 물센터의 거대억새 생산력 검정을 위한 시험 포장에서 수행 되었다. 시험포장에는 거대 1호, 억새 2호, 우람억새, 도입 종(3배체)억새 및 일반 물억새의 5종류의 억새를 4년간 재 배 중이며, 각 억새종별 시험포장의 면적은 10 × 10 m로 난 괴법 3반복으로 구성되어 있다. 이 중에서 거대 1호를 대상 으로 이화명나방 유충의 발생형태를 조사하였다.
본 연구를 위한 이화명나방 유충의 조사기간은 이화명나 방의 2화기 후 2012년 10월경부터 이듬해 2013년 3월경까 지이며 월 1회 조사를 수행하였다. 조사방법으로는 매월 거 대 1호 시험포장에서 1 x 1 m 내의 식물체 지상부(줄기)를 지상에서 최대한 근접하게 절단하여 수확하였고, 수확된 식 물체 개체에 해당하는 지하부(뿌리)를 채취하였다. 수확된 지상부는 초장 및 줄기수 조사 등의 생육조사를 행하였으며 생육조사가 완료된 줄기는 종으로 쪼개어 이화명나방 유충 의 발생여부(유/무)를 조사하였다. 또한 기온에 따른 이화명 나방 유충의 이동형태를 알아보기 위하여 월별로 절단된 줄 기 최하단 부분으로부터 유충이 출현된 위치까지의 높이를 측정하였다. 이와 동시에 각 조사지점에서 채취한 지하부도 종으로 쪼개어 이화명나방 유충의 밀도를 조사하였다.
거대억새를 가해하는 이화명나방에 의한 피해정도를 분 석하기 위하여 억새의 생육말기인 2012년 11월경에 각 시 험구내의 단위면적당(1 x 1 m) 거대억새 전체 마디수(경수) 를 조사하였으며 이화명나방에 의해 줄기에 구멍이 뚫어져 있거나 부러진 마디를 피해를 입은 식물체로 판단하여 그 경수를 조사하였다.
결과 및 고찰
억새에 기주하는 이화명나방의 2화기 이후 유충의 월동 양상을 분석하기 위하여 억새 식물체 지상부 및 지하부 내 의 밀도를 조사한 2012년 10월부터 2013년 3월까지의 전남 무안군의 각 월별 평균기온을 살펴보면 2012년 10월은 14.6°C, 11월은 7.6°C, 12월은 0.4°C이었으며, 이듬해 2013 년 1월의 월평균기온은 -0.5°C, 2월에는 1.2°C, 3월에는 5. 4°C이었던 것으로 나타났다(Fig. 1).
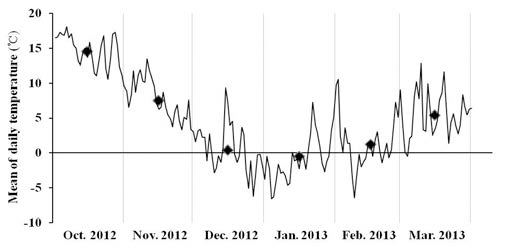
Fig. 1.
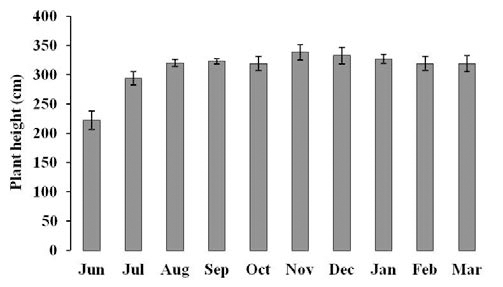
Mean of Daily temperature from October 2012 to March 2013 in Bioenergy Crop Research Center, RDA located in Muan, Jellanamdo in Korea. Diamond marks indicate the mean of monthly temperature.2012년 6월부터 2013년 3월까지 기주식물인 거대 1호의 초장을 조사한 결과, 억새 생육초기인 6월에는 평균 223 cm 이었으며 7월에는 294.3 cm로 초장의 증가폭이 가장 컸다 (Fig. 2). 8월 이후부터 이듬해 3월까지는 초장의 증가폭은 크지 않은 것으로 나타났으며 평균 초장 300 cm 이상이었다.
거대억새에 기주하는 이화명나방 유충의 밀도를 조사한 결과는 Fig. 3에 나타내었다. 2012년 10월에 거대억새 지상 부에서 출현한 이화명나방 유충의 수는 평균 23마리이었으 며 11월에는 4마리이었고 12월과 이듬해 2013년 1월에는 각각 1마리씩 출현하였다. 하지만 그 이후 2월과 3월에는 거대억새 지상부에서 이화명나방 유충이 발견되지 않았다 (Fig. 3a). 유충이 발견된 줄기의 발생위치별 유충수를 월별 로 분석한 결과, 가장 많은 유충이 발견된 10월의 경우에는 수확한 거대억새 지상부의 최하단으로부터 21~25 cm부근 에서 12마리가 발견되어 가장 높은 밀도를 나타내었으며 그 다음으로는 10~15 cm 부근에서 5마리, 16~20 cm와 26~30 cm 부근에서는 각각 3마리씩이 발견되었다. 11월경에는 10~15 cm부근과 16~20 cm 부근의 거대억새 줄기 하단부위에서 각각 2마리씩 발견되었다. 또한 12월경과 이듬해 2013년 1 월경에 1마리씩 발견된 유충의 서식위치는 10~15 cm 부근 인 것으로 나타났다.
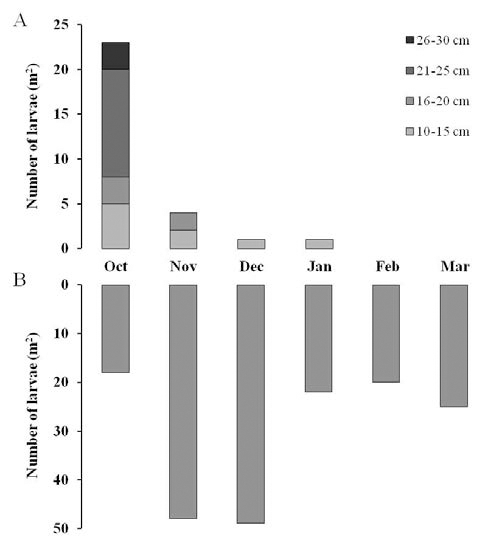
Fig. 3.
Numbers of larvae of Chilo suppressalis observed on stems (A) and rhizomes (B) of Miscanthus sacchariflorus cv. Geodae 1.더 나아가 10월부터 이듬해 3월까지 거대억새 지하부(뿌 리)에 기주하는 이화명나방 유충의 출현밀도를 조사한 결 과, 10월경에는 평균 약 18마리가 관찰되었으나 그 이후 11 월과 12월에는 각각 48마리, 49마리가 관찰되며 가장 높은 밀도를 나타내고 있음을 알 수 있었다(Fig. 3b). 그리고 1월, 2월, 3월에는 약 20~25마리의 밀도를 유지하고 있었다. 이 처럼 기온이 하강함에 따라 지상부의 이화명나방 유충의 밀 도는 감소하고 그와 동시에 지하부(뿌리) 내의 유충수가 증 가하는 결과로부터 이화명나방의 유충은 기온이 10°C이하 로 내려가는 11월 전후로 뿌리 가까운 줄기로 이동하거나 줄기 기부 및 지하부(뿌리)로 이동하여 월동을 하는 것으로 확인되었다.
거대억새 지하부의 유충조사를 행한 결과, 흥미롭게도 줄 기 기부 뿐만 아니라 억새 뿌리(지하경) 내에도 유충이 다 수 출현하는 것을 확인할 수 있었다. Fig. 4a의 경우는 줄기 기부 약 5 cm 아래에서 월동 중인 유충을 나타내고 있으며, Fig. 4b에서 보여지듯이 뿌리 내부에서도 월동 중인 유충을 발견할 수 있었다. 일반적으로 이화명나방 유충은 벼의 수 확이 이루어진 후 그 대부분이 벼의 그루터기, 줄기 및 논 가장자리의 잡초로 이동하여 월동을 한다고 알려져 있다 (Atapour & Moharramipour, 2009; John et al., 2014; Yu, 1980). 하지만 본 연구결과, 억새에 기주하는 이화명나방 유충의 월동장소는 줄기 기부 또는 지하경과 같이 지하부위 라는 점이 현재까지 알려진 벼에서 기주하는 이화명나방의 유충과는 다른 월동형태를 보이는 것으로 최초 확인되었다.
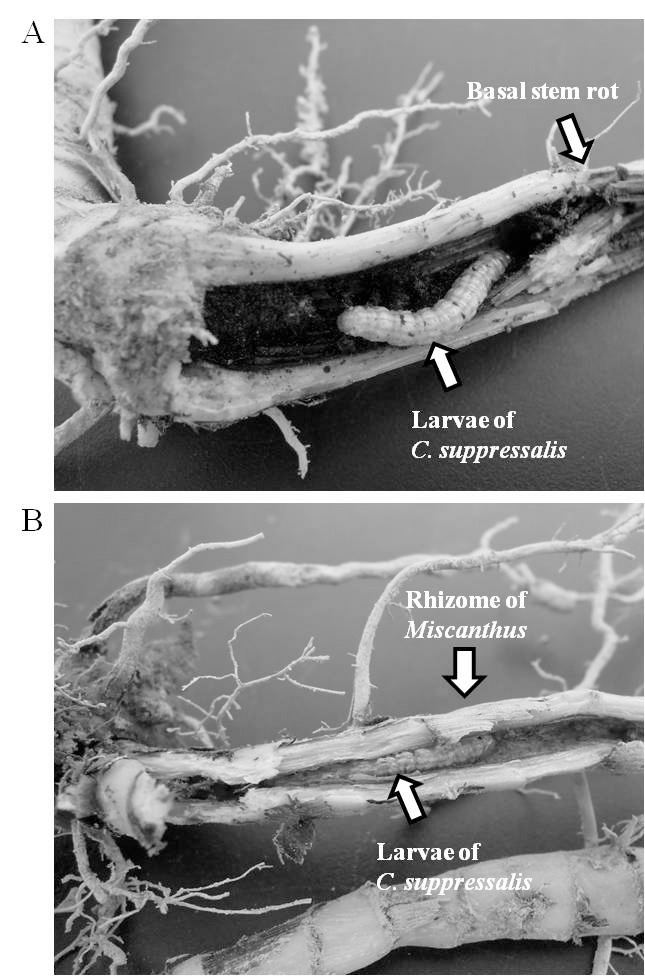
Fig. 4.
Overwintering locations of Chilo suppressalis larvae on basal stem rot (A) and into the rhizome of Miscanthus sacchariflorus cv. Geodae 1 (B).억새류 중에서 거대억새 지하경의 가장 큰 특징 중의 하 나는 다른 참억새 및 일반 물억새와 비교하여 지하경의 굵 기가 2배 이상 굵으며 억새 전체 지하경의 약 90% 이상이 지하 30 cm 이내 범위에 분포한다고 알려져 있으며(An et al., 2012; Mann et al., 2013), 또한 지하경과 줄기의 단면 을 보면 꽉 차있지 않고 가운데에 공간이 존재한다(Fig. 5). Choi et al. (1979)에 의하면 이화명나방의 산란선호성은 기 주식물 줄기 굵기 등의 형태적 특성과 유의한 상관관계가 있다고 보고한 바 있다. 이처럼 거대억새의 굵은 줄기 및 지 하경의 특성은 이화명나방의 유충이 외부로부터 인위적인 방제의 행위를 피하는 동시에 월동하기에 최적의 장소이었 음을 시사한다.
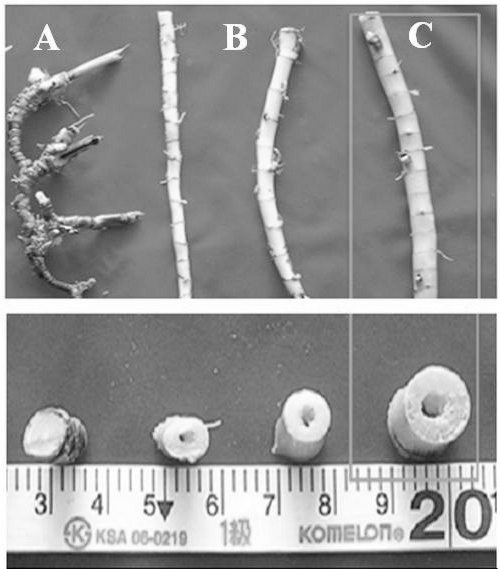
Fig. 5.
Rhizomes of Miscanthus spp. (A, Miscanthus sinensis; B, common Miscanthus sacchariflorus; C, M. sacchariflorus cv. Geodae 1).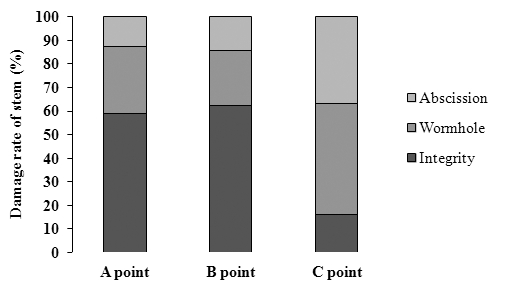
Fig. 6.
Damage rates to stems of Miscanthus sacchariflorus cv. Geodae 1 grown at three points by Chilo suppressalis.이화명나방 유충의 가해로 인한 거대억새의 피해정도를 조사하기 위하여 포장내 3개의 지점을 대상으로(A, B, C지 점) 1 x 1 m 내의 전체 경수를 조사하였으며 그 중 피해를 입 은 줄기는 이화명나방에 의하여 구멍이 뚫린 줄기(wormhole) 와 부러져 있는 줄기(abscission)로 나누어 조사하였다. 그 결과, A지점, B지점 및 C지점에서의 거대억새 전체 평균 줄기수는 각각 80.5, 83.5, 70.5개 이었다(Fig. 5). A지점에 서의 전체 줄기 중 구멍이 뚫린 줄기 비율은 전체의 28.6% 이었으며, 부러진 줄기수는 전체의 12.4%이었다. 이화명나 방에 의한 피해율은 A, B지점에서 50% 이하로 피해가 경 미한 상태였으나, C지점에서는 구멍이 뚫린 줄기비율이 전 체의 46.8%로 높은 편이었으며, 부러진 줄기비율도 36.9% 로 나타나 이화명나방에 의한 피해줄기 비율이 83.7%인 것 으로 조사되었다.
본 연구 결과, 1970년대 국내 대표적인 벼 가해 해충으로 잘 알려진 이화명나방이 섬유질계 바이오에너지작물인 억 새를 대규모 식재 및 재배하였을 시 바이오매스 생산성에 악영향을 미칠 수 있는 잠재적 해충임을 알 수 있었다. 보다 심각하게 생각되는 부분은 바이오매스용 억새의 지상부 수 확시기가 2~3월경인데 그 시기에 이화명나방 유충 대부분 은 억새의 밑둥 줄기 및 지하경 내에서 월동하고 있다는 점 을 볼 때 수확에 의한 유충 밀도감소 등의 재배적 방제방법 효과가 거의 없을 것으로 판단된다. 특히 벼에서 2화기 이 후 볏짚 또는 벼그루터기에 기주하며 월동하는 양상과 전혀 다르게 억새밭에서의 월동위치가 지하 5~10 cm 이하인 억 새 줄기기부와 지하경 내에까지 분포한다는 것을 감안할 때 지금까지 알려진 방제방법(불태우기, 화학적 방제)의 효과 도 거의 없으리라 추측된다. 이러한 연구결과를 바탕으로 금후에는 억새밭에서의 새로운 방제방법(화학적, 재배적, 생물학적)에 대한 연구개발이 지속적으로 이루어져야 할 것 이다.
적 요
본 연구는 바이오에너지 원료생산을 위해 대규모로 억새 를 재배할 시에 억새의 생육 및 수량감소에 영향을 미치는 잠재적 유해요인을 사전에 대비하며 효과적으로 억제할 수 있는 방법 개발을 위한 기초자료로 활용하고자 수행하였다. 전남 무안의 억새 재배지에서 2012년 10월부터 이듬해 3월 까지 이화명나방 유충의 월동양상 및 밀도를 조사하기 위해 월별로 유충의 출현 위치, 가해 피해양상을 조사하였으며 주요 결과를 요약하면 다음과 같다. 섬유질계 바이오에너지 작물인 거대억새에 기주하는 이화명나방 유충밀도를 2화기 이후인 2012년 10월에서 2013년 3월까지 조사한 결과, 월 평균 기온이 내려갈수록 유충의 출현위치가 점차 지상부의 아래부위 또는 줄기기부로 이동하는 양상을 확인하였으며 월평균 기온이 10°C이하로 내려가는 11월 및 12월경에는 지하경(뿌리) 부위에서 가장 많은 이화명나방 유충 밀도를 나타내는 것으로 조사되었다. 억새밭에서 이화명나방 피해 는 줄기 침입구멍 존재여부와 부러진 줄기로 판단하였는데 전체 줄기 중 구멍이 뚫린 줄기 비율이 28.6%이었으며, 부 러진 줄기수는 전체의 12.4%이었으나 심한 곳은 구멍뚫린 줄기비율이 46.8%, 부러진 줄기비율도 36.9%로 나타났다. 억새밭에 기생하는 이화명나방 유충의 경우, 지하 5cm 이 하의 줄기기부 심지어는 뿌리내부에서 월동을 한다는 것을 확인하였으며 이로 인해 일반적인 방제방법으로는 이화명 나방 유충의 밀도를 감소시킬 수 없을 것으로 판단되어 지 속적으로 억새밭 이화명나방 방제방법의 개발이 개발되어 야 한다.
