서 론
재료 및 방법
시험 재료
재배 방법 및 농업 특성 조사
기상 환경 조사
통계 분석
결과 및 고찰
최근 2년간 밀 파종기부터 성숙기까지 기상과 생육 특성 변화
출수기 및 성숙기 변화 분석
간장 및 도복 변화 분석
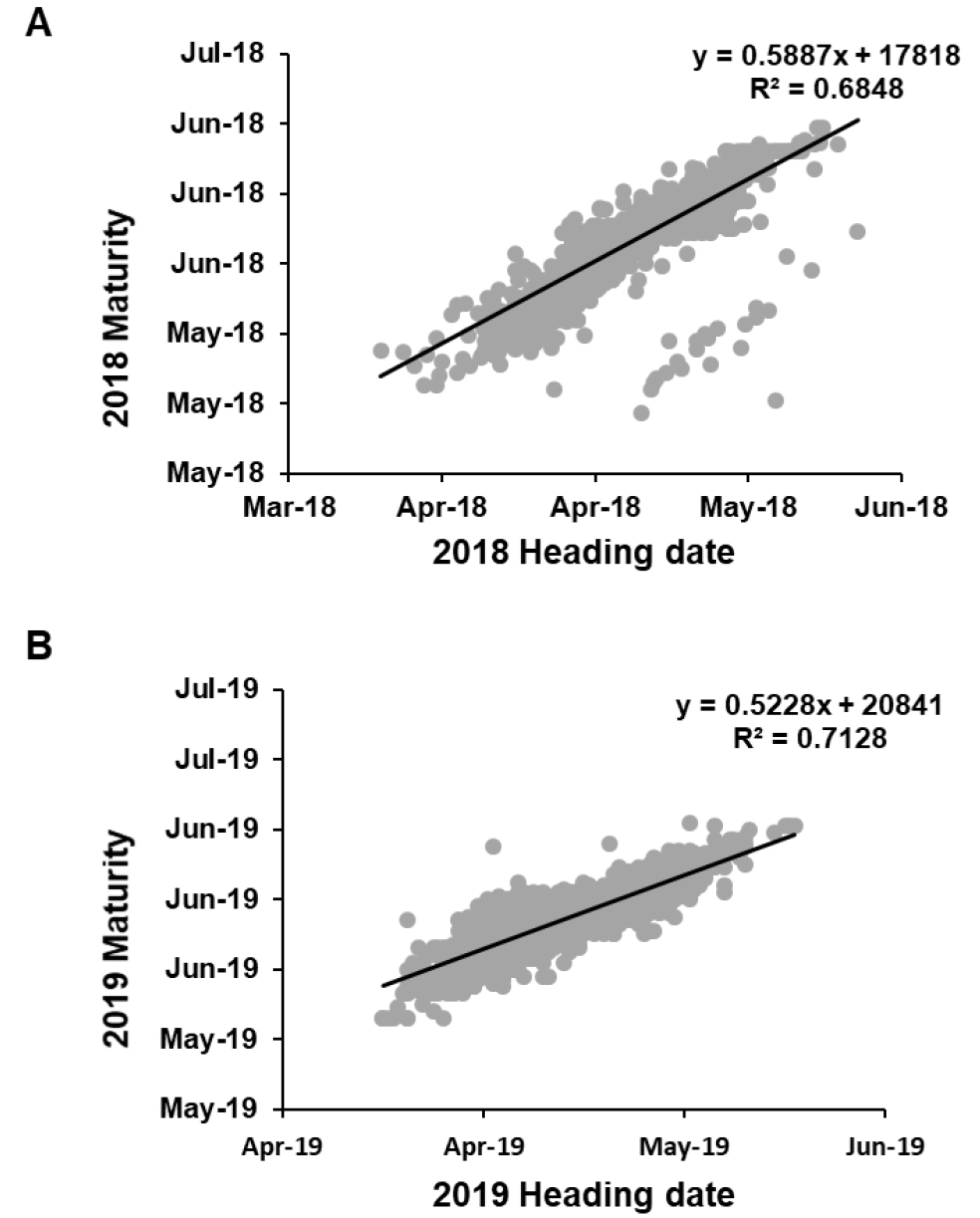
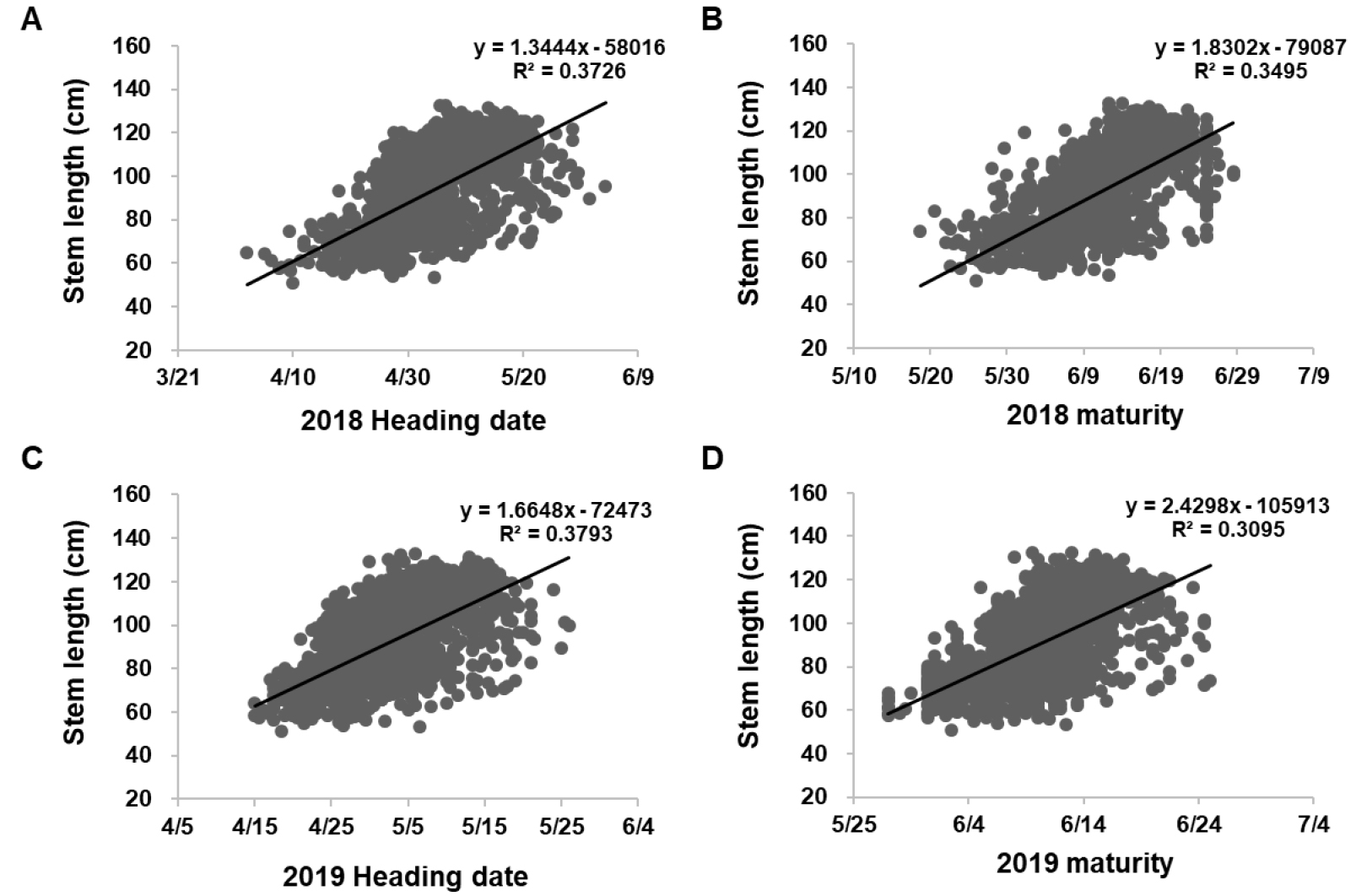
간장과 출수 및 성숙과의 관계
적 요
Supplement
서 론
밀은 벼, 옥수수 등 주요 식량작물 중 하나로서 전 세계적으로 매우 중요한 작물이다(Howell et al., 1995). 밀의 주요 농업적 특성은 초장, 분얼 수, 엽수 등 식물체 특성과 종자의 크기, 색, 모양과 같은 종자의 특성을 포함하고 있다. 이와 함께 출수기와 성숙기는 밀의 주요 농업특성과 더불어 제외할 수 없는 중요한 요인이다. 종자의 모양이나 종자 수는 밀의 수량성과 매우 밀접하게 관련되어 있다(Fischer et al, 1984; Sinclair & Jamieson, 2008). 따라서 밀의 수량은 단위 면적당 종자 수 또는 크기에 따라 변화될 수 있다(Fischer, 1984; Calderine & Slafer, 1998; Slafer & Araus 2007; Fischer & Edmeades, 2010). 그러나 국내 밀 육종은 밀의 종자 특성뿐만 아니라 국내 농업체계와 기후조건에 적합한 밀 품종 육성을 위해 출수기와 성숙기를 앞당겨 빠른 수확으로 이어질 수 있도록 수행되어 왔다(Son et al, 2017). 최근 분자육종의 발전과 함께 밀의 주요 농업특성과 직접적 연관이 되는 유전자 마커(Rht, Ppd-1, Vrn-1 등)가 개발되었고, 이들 유전자는 국내 품종 및 주요 자원에서 평가되었다(Shin et al, 2014). 그러나 앞으로 국내 환경에 적합한 뛰어난 품종 개발을 위해서는 세계 곳곳의 많은 자원들에 대한 수집과 평가가 수행되어야 한다. 세계적으로도 짧은 기간에 성숙하여 고온이 오기 전에 수확을 하기 위해 출수와 개화가 빠른 일장둔감형 밀의 특성을 갖는 밀을 육종하고 있다. 파성이 충족되었을 때, 일장에 상관없이 기온 상승에 의해 생식생장 단계로 넘어가기 때문에 밀의 재배 기간을 단축 시킬 수 있다(Guedira et al, 2016).
기후는 작물의 생리와 생장, 발육 등에 관련된 작물의 근본적인 과정에 영향을 미치기 때문에 농업에 있어 기후변화는 매우 중요한 요인이다(Meza & Silva, 2009). 따라서 기후변동성의 효과 및 기후변화에 대한 작물 생리 및 생산성 등이 포괄적으로 연구되었다(Parry et al, 2004). 최근 고온 현상과 이상기온에 따른 밀의 생육이 문제가 되고 있다. 기온과 강수량과 같은 환경 변화는 밀의 개화 및 생산량과 매우 밀접하게 연관되어 있다(Sommer et al, 2013). 특히, 개화는 종자의 수를 결정하는 가장 중요한 특징이며, 이 시기의 가뭄, 급격한 고온이나 저온 현상은 개화에 심각한 영향을 미칠 수 있다.
따라서 기후변화에 대하여 적응성이 크거나 둔감한 밀 계통 또는 품종의 개발이 필요하다. 본 연구는 연차 간 기후변화에 따른 유전자원의 실제 출수기와 성숙기 등 농업특성을 비교 분석하여 육종에 한국의 기후 변화에 적합한 밀 품종을 개발하는데 도움이 되는 자료를 제공하고자 수행 하였다.
재료 및 방법
시험 재료
본 연구에 사용된 1969개 밀 유전자원은 농촌진흥청 유전자원센터로부터 분양 받은 것으로서 각각 한국에서 육성된 계통 및 재래종 밀을 포함하여, 일본, 멕시토, 러시아, 미국 등 세계 60 여 개 국의 품종 및 계통으로 구성되어 있다. 1969개 밀 유전자원 중에는 국내 재래종 및 육성 계통 및 멕시코 자원이 342점과 367점으로 가장 많이 포함되어 있으며, 중국과 미국자원이 150여점씩, 아프가니스탄 및 에티오피아 자원이 70여점씩 포함되어 있으며, 153점은 원산지가 불분명 하였다(Fig. 1D). 그리고, 그 중 발아 및 생육이 이루어지지 않은 자원 29개를 제외하고, 1940개 밀 유전자원의 출수기, 성숙기, 간장, 수장 및 도복 등 주요 농업특성을 2018년부터 2019년까지 2회 조사하였다(Fig. 1A, 1B).
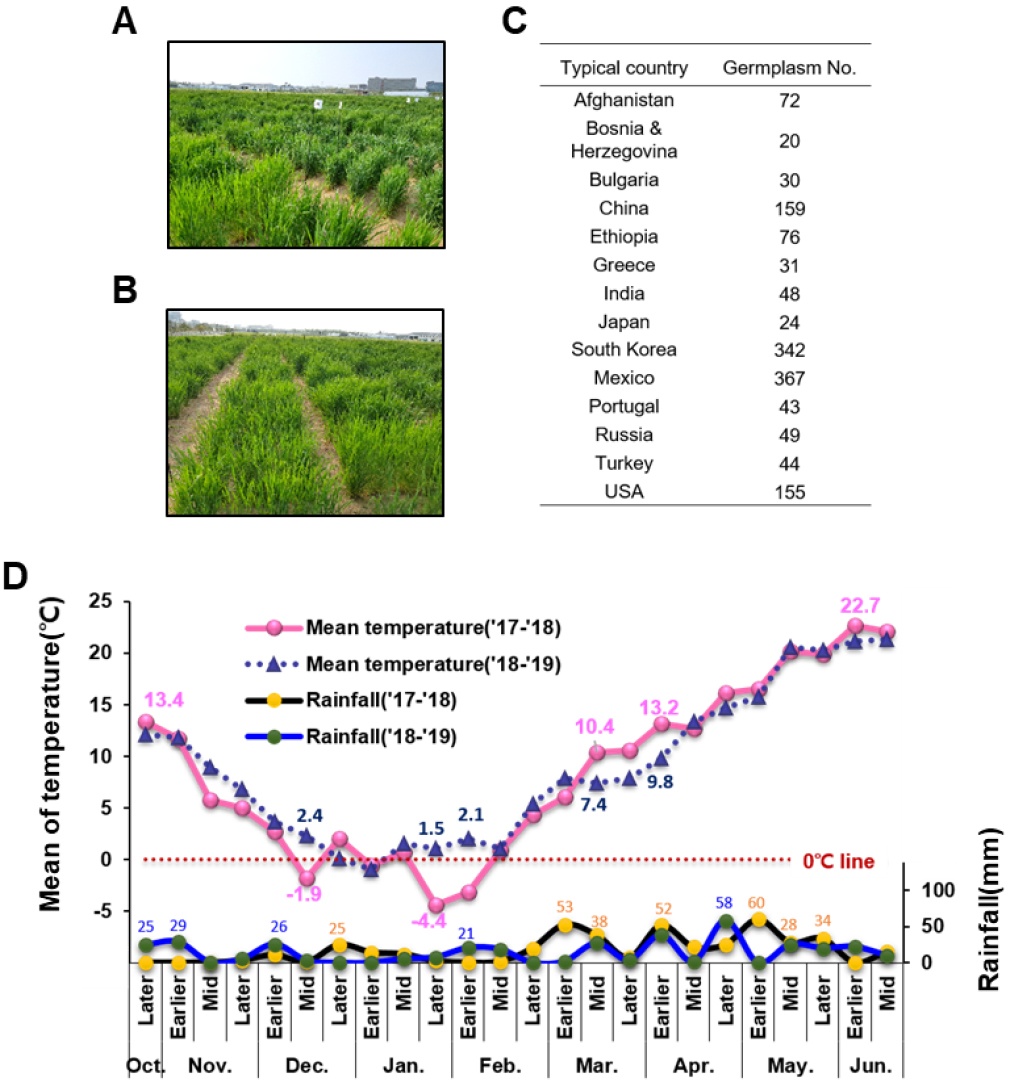
Fig. 1
The 1969 wheat germplasm were collected from about 60 countries and typical phenotypic traits wheat genetic resources were estimated. A and B; The picture shows 1969 wheat germplasm grown at wheat breeding fields of National Institute of Crop Science (NICS), Wanju in Korea in 2018 and 2019. C; Typical origin countries and number of germplasm were list up. D; The graph shows average temperature and raining for two years (2018. Jan.~Apr. and 2019. Jan.~Apr.).
재배 방법 및 농업 특성 조사
2018년부터 2019년 2년 동안 농촌진흥청 국립식량과학원 전작 포장에서 재식 밀도를 0.4 m × 6 m로 10월 25일에 파종하였다. 시비 및 재배는 농촌진흥청 표준재배법에 따라 N2 : P2O5 : K2O = 7.9 : 7.4 : 3.9 kg/10a을 기준으로 실시하였고, 수확은 6월 중순에 실시하였다. 농업 특성은 밀의 간장, 수장, 출수기 및 성숙기와 쓰러짐 등 특성 조사는 농촌진흥청 농업 과학 기술 조사 분석 기준(RDA, 2012)에 준하였다. 밀 유전자원 종피의 색도는 색차계(Minolta JS-555, Minolta Camera Co., Ltd, Japan)를 이용하여 분석하였다.
기상 환경 조사
기상 환경은 2017년부터 2019년 10월 파종기부터 수확기인 이듬해 6월까지 9개월의 전주시 기온과 강수량 자료를 기상청으로부터 자료를 다운받아 본 연구에 활용하였다. 기온과 강수량은 각각 년도의 평균으로 비교하여 분석하였다.
통계 분석
시험성적은 R 프로그램(R-3.5.1)으로 피어슨의 상관계수를 이용하여 상관분석(n=3)을 실시하였고, 통계적 유의 수준은 P<0.05 이다.
결과 및 고찰
최근 2년간 밀 파종기부터 성숙기까지 기상과 생육 특성 변화
최근 국내 밀 재배 시기의 기후변화로 인해 밀과 같은 동계작물의 생육 변화가 초래하고 있다. 국립식량과학원 전작포장에서 약 60여 개국 2000여점의 밀 유전자원을 가지고 2년간(2017~2018과 2018~2019년) 기후변화에 따른 밀의 생육변화를 관찰하였다(Fig. 1A, 1B, 1D).
두 연도에서 기상변화를 확인하고자 기상청 자료를 바탕으로 밀 유전자원을 파종 후부터 성숙기(6월 중순)까지의 기온 및 강수량을 분석한 결과 파종일(10월 중순)부터 성숙기(6월 중순)까지 평균기온 변화를 비교했을 때, 2017~2018 (이후 2018년이라 호칭)년과 2018~2019 (이후 2019년이라 호칭)년의 평균온도는 각각 8.6°C와 9.0°C로 0.4°C로 통계적으로 유의한 차이를 보이지 않았다(Fig. 1D). 그러나 밀 생육단계별 기온 변화는 두 연도 간에 차이가 컸다. 두 해의 파종 후부터 12월까지 평균온도는 각각 5.5°C와 6.5°C로 2019년(2018년파종)이 1°C 이상 높았다. 또한 1월부터 2월까지 밀의 생육이 정지된 시기의 평균온도는 각각 -0.4°C와 1.7°C로 2019년이 약 1.3°C 높았고, 특히 2019년도는 1월 초를 제외하고 2월 말까지 약 2개월간 평균온도가 영상 기온을 유지하였다. 이 온도변화에 따라 생육이 정지되는 시기인 월동기에 밀의 초기 생육 및 길이 생장이 있었을 것으로 예상된다. 국립식량과학원에서 매년 국내 대표 밀 품종의 생육 결과를 보고하는 ‘맥류 작황 연구보고서’에 따르면 생육재생기는 2월 5일로써 2019년도는 2월 7일로 2018년도 보다 5일 빨랐으며, 초장은 평균 7 cm로서 2018년도 6 cm보다 1 cm 긴 것으로 확인되었다. 2019년 지상부 건물중은 평년보다 1.8배 증가하였고, 2018년도와 비교하여 1.5배 증가한 것으로 확인되었다(data not shown).
3월부터 6월 중순까지 밀 전체 생육기간의 평균기온은 각각 15.5°C와 14.5°C로 2018년이 약 0.9°C 높다. 밀의 생육시기별 두 연도간 온도변화를 살펴보면, 2018년과 2019년도 영양생장기(3월초~4월중순) 평균기온이 각각 10.6°C와 9.3°C로 2018년이 약 1.3°C 높았으며, 특별히 3월 중순부터 4월 중순까지는 약 2.0°C 가량 높았다. 그리고 밀의 출수 후 종자 등숙 및 성숙기인 5월 초부터 6월 중순까지 평균기온은 각각 20.3°C와 19.8°C로 거의 차이를 보이지 않았다.
2017~2018년과 2018~2019년의 강수량 변화를 기상데이터를 통해 확인해 본 결과 파종 후 성숙기까지 평균 강수량은 2018년도에 각각 17.6 mm로써 2019년도의 14.6 mm보다 약 3 mm 정도 더 많았다(Fig. 1D). 밀 생육시기별 강수량 변화는 파종 후부터 12월까지 평균강수량이 각각 5.7 mm와 12.6 mm로 2019년(2018년파종)에 6.9 mm 이상 더 많았다. 밀 생육정지기간인 1월부터 2월까지의 평균강수량은 각각 8.3 mm와 8.7mm로 거의 비슷하였다. 밀 생육 전 기간인 3월부터 6월 중순까지 평균강수량은 각각 30.3 mm와 19.0 mm로 2018년이 약 11.3 mm 이상 강수량이 많았다. 밀 생육기간을 더욱 세분화 하여 본 결과 밀의 영양생장기인 3월부터 4월 중순까지 평균강수량 또한 각각 34.3 mm와 14.6 mm로 2018년이 약 19.7 mm 이상 많았다. 3월 중순부터 4월 중순까지는 2018년의 평균 강수량이 약 11.9 mm 이상 많았다. 그리고 밀의 성숙시기로 들어가는 5월부터 6월 중순까지 평균강수량 또한 각각 27.4 mm와 15.5 mm로 약 10 mm 이상 차이를 나타냈다.
기온과 강수량을 종합하여 볼 때, 밀의 생육시기별 기온과 강수량을 비교하면, 2018년도의 경우 파종(2017년 10월)부터 다음해 2월(2018년)까지 기온과 강수량이 상대적으로 낮았지만, 3월 중순부터 4월 중순까지 평균기온과 강수량이 각각 2°C, 11.9 mm 이상 높았다. 특히 2월 말부터 3월 중순까지 평균 강수량은 2018년도 때 약 27.2 mm 이상 많은 특징을 보였다. 밀의 재생기인 2월부터 영양생장이 시작되는 시기에 2019년도의 강수량이 매우 낮았고 기온이 높았다는 것을 알 수 있다.
출수기 및 성숙기 변화 분석
밀의 성숙기는 고온다습한 국내 초여름 기후에 따라 등숙에 불리한 재배환경과 아울러 이모작 작물 준비 관계로 6월 10일 이전에 수확이 가능하도록 품종을 육성해 왔다. 국내 환경에서 밀 성숙기는 출수기와 높은 상관관계가 있다(Supplementary Fig. 1). 온도, 파성(vernalization), 일장(photoperiod)은 밀의 숙기와 밀접하게 관련되어 있는 것으로 여러 연구를 통해 보고되어 있다(Law et al, 1976). 특히 밀의 발달과 개화는 온도와 일장에 매우 민감하다(Baker et al, 2006). 2018년도 출수기와 성숙기의 상관관계보다 2019년 출수기와 성숙기의 상관관계의 유의성이 더욱 컸다. 이는 영양생장기인 2018년 2018년 3월에서 4월 초에 평년 온도보다 높은 기온에 따라 출수기의 변화로 인한 것으로 여겨졌다(Fig. 1D).
2018년과 2019년도 평균 출수기는 각각 5월 3일, 5월 2일로 비슷하게 확인되었고, 성숙기는 6월 11일로 같았다(Fig. 2A). 수원242호(한국)의 출수기는 2018년과 2019년도 4월 2일과 4월 17일로 가장 빨랐고, Cr9349 (네델란드)는 2018년 5월 31일로 가장 늦게 출수가 되었으며, 2019년도에는 April Bearded (영국) 자원이 5월 26일로 가장 늦었고, Cr9349와 Bearded Red (영국)이 5월 25일로 확인되었다. 성숙기는 수원303호가 2018년 5월 22일, 2019년 5월 28일로 가장 빨랐고, April Bearded와 Bearded Red는 2018년 6월 28일로 가장 늦었지만, 2019년은 MK2565 (멕시코)가 6월 25일로 가장 늦었고, 두 자원은 6월 24일로 확인되었다(data not shown).
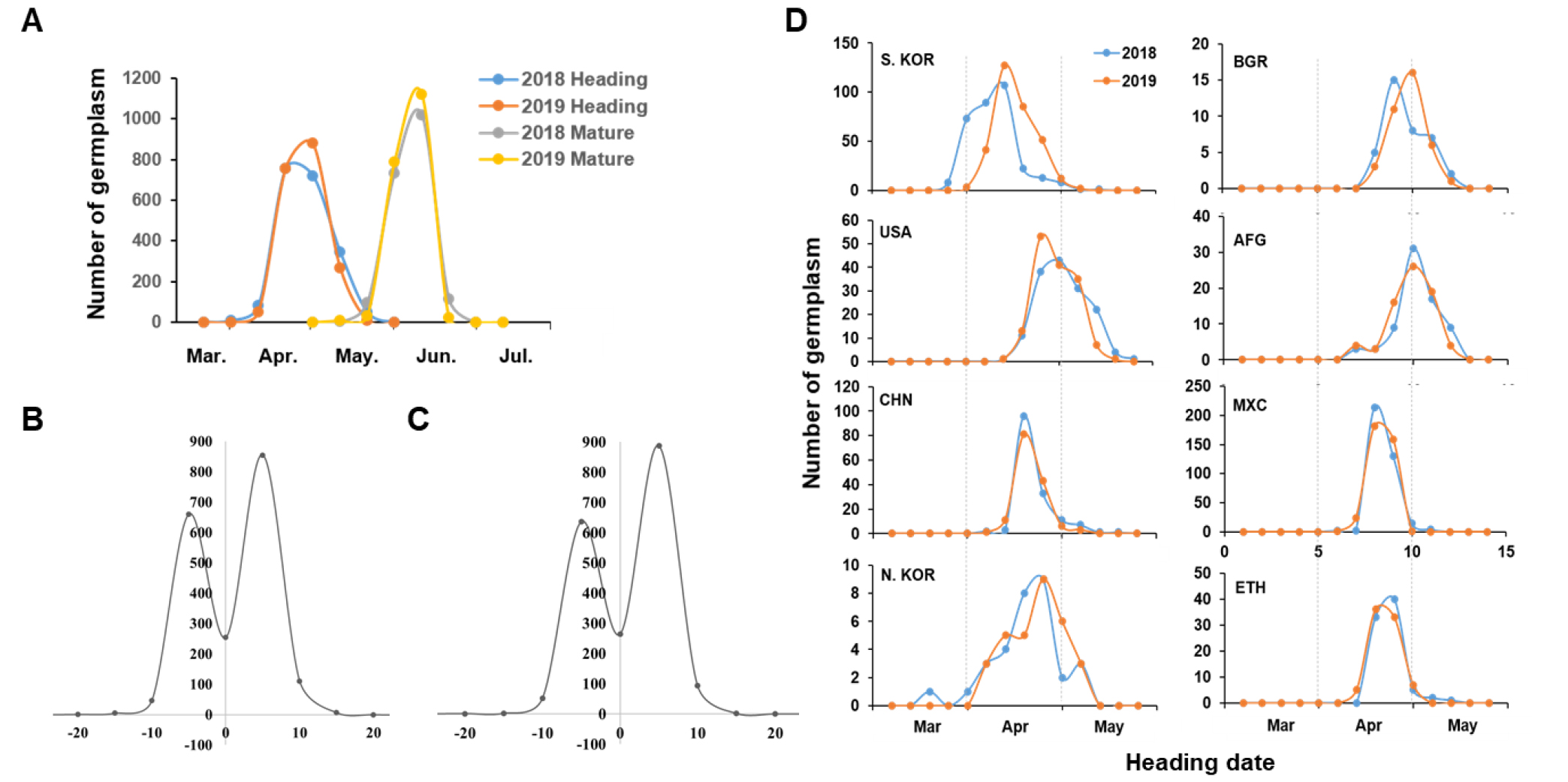
Fig. 2
Heading date and maturity of 1969 wheat germplasm were estimated and compared by typical origin countries of genetic resources. A; Number of resources in heading date and maturity, respectively, B; Changed number of resources in the heading date in 2019, C; Changed number of resources in the maturity in 2019, D; Heading and mature dates for two years were divided by typical origin countries.
출수기와 성숙기는 2018년과 20019년 차이가 있는 것으로 확인되었다(Fig. 2A). 2018년에 비해 출수기가 늦어진 자원은 713개, 빨라진 자원은 973개, 그리고 변화가 없는 자원은 254개로 확인되었다(Fig. 2B). 2018년도 대비 출수기가 늦어진 대표 자원은 국내 재래종밀 중 한 자원으로써 17일이나 차이가 있었다. 11에서 15일 정도 늦어진 자원은 5개, 6-10일 늦어진 자원은 47개, 1-5일 늦어진 자원은 660개로 확인되었다. 이와 반대로 출수기가 빨라진 자원 중 11일 이상 빨라진 자원은 7개였고, CI7717 (러시아)과 Thurorato-1은 13일로 가장 빨랐다. 6-10일 빨라진 자원은 111개, 1-5일 빨라진 자원은 855개로 확인되었다. 국가별로 한국, 북한, 불가리아의 밀 자원들은 2019년이 2018년에 비해 출수기가 대부분 늦어지는 경향을 보였다. 특별히 국내 자원의 출수기는 다른 주요 자원국 밀에 비해 평균 10에서 20일가량 빨랐으며, 2019년 출수도 10일가량 큰 폭으로 늦어졌다. 반대로 미국의 자원들은 빨라지는 경향을 나타냈다. 그리고, 중국, 아프카니스탄, 멕시코, 에티오피아의 자원들은 2018년과 2019년도 출수기가 거의 비슷하였다.
2018년에 비해 성숙기가 늦어진 자원은 692개, 빨라진 자원은 984개, 그리고 변화가 없는 자원은 264개였다(Fig. 2B). 그 중 D.I.V.6694가 16일 이상으로 가장 늦었다. 그 다음으로 Mk2565 (멕시코)가 13일, S1819 (인도)가 12일로 늦었다. 6-10일 성숙기가 늦어진 자원은 52개, 1-5일 늦어진 자원은 637개였다. 이와 반대로 Hezohegyesi 7286 (헝가리)와 Thuroato-1은 11일이상 성숙기가 빨라진 것으로 확인되었다. 그 다음으로 6-10일 빨라진 자원은 94개였고, 1-5일 빨라진 자원은 888개였다. 2년간 재배한 유전자원 중에서 출수기와 성숙기의 변화가 없는 자원은 수원211 등 37개 자원이었다. 이들 자원은 2년 동안의 환경변화에 관계 없이 같은 기간에 출수와 성숙을 하는 것으로 나타났다. 국가별로 나누었을 때, 수원211 등 한국은 7개, 아프카니스탄 4개, 오스트레일리아 1개, 중국 3개, 에콰도르 1개, 이라크 2개, 레바논 1개, 마케도니아 2개, 멕시코 7개, 포르투갈 1개, 러시아 1개, 터키 1개, 미국 2개 자원과, 원산지가 불확실한 자원이 4개였다. 그리고 출수기는 변화가 없었지만, 성숙기는 늦어진 자원은 114개였고, 대부분 1일에서 3일 정도 성숙기가 늦어졌다. 성숙기만 빨라진 자원은 103개였고, 성숙기 또한 1일에서 3일 정도 성숙기가 늦어졌다. 성숙기는 변화가 없고 출수기가 늦어진 자원은 120개였고, 빨라진 자원은 107개로 확인되었다. 이 두 결과 또한 대부분이 1일에서 3일간의 차이가 가장 많았다(data not shown).
출수에서 성숙기까지 일수(등숙일수)가 가장 짧은 자원은 Pyrothryx (헝가리)와 CI7717 (러시아)로 18년도 25일, 19년도 32일로 19년도 등숙일수가 7일이 더 길어졌다. 18년도 대비 등숙일수가 가장 늘어난 자원은 D.I.V.6694 (아르헨티나)로 18년도 대비 19년도에 등숙을 위한 시간이 15일이 더 소요되었다. 세부적으로 10일 이상 더 소요된 자원은 6개, 5-9일은 101개, 1-4일은 727개로 확인되었고, 반대로 1-4일 이하로 소요일수가 줄어든 자원은 709개, 5-9일은 94개, 10일 이상 줄어든 자원은 9개로, 그 중 수원242호(한국)는 2019년 성숙일수가 31일로 18년도 대비 16일이 감소되었다. 그리고, 등숙일수에 변화가 없는 자원은 bearded Red 등 294개 자원이 확인되었다(data not shown). 또한 종자의 색은 성숙기, 간장, 수장과는 음의 상관관계를 나타냈고, 출수기와는 양이 상관을 나타냈다. 특히 출수기는 붉은색 계열이 0.115, 노란색 계열이 0.125로 확인되었고, 성숙기는 각각 -0.199와 -0.246으로 확인되었다. 이러한 결과는 색이 붉은색과 노란색 계열의 유색의 특징을 가질 수록 출수기는 느려지지만, 반대로 성숙기는 빨라지기 때문에 밀의 전체적인 생육기간은 빨라질 수 있는 것을 의미할 수 있다. 간장과 수장 또한 종자의 색이 진할 수록 감소하는 경향을 나타냈다(Table 1).
Table 1.
The correlation of grain color with agriculture characteristics of wheat.
| Heading date | Maturity | Stem length (cm) | Spike length (cm) | |
| L (Lightness) | 0.091 | -0.214 | -0.084 | -0.089 |
| A (Redness) | 0.115 | -0.199 | -0.051 | -0.097 |
| B (Yellowness) | 0.125 | -0.246 | -0.078 | -0.117 |
간장 및 도복 변화 분석
파종 후 출현부터 성숙기까지의 온도변화는 밀의 모든 생장 단계에 큰 영향을 미친다(Slafer & Rawson, 1994). 2년간 밀 유전자원의 전반적인 출수 및 성숙기 변화뿐만 아니라 이삭 및 망장(까락의 길이)의 길이에는 변화가 없었다(Fig. 1A, 3C, 3D). 하지만 2019년 성숙기에 조사한 유전자원의 간장은 2018년과 비교하여 평균 약 20cm 가량 더 증가한 것으로 확인되었다(Fig. 3A, 3B). 2019년에 간장이 증가한 이유에 관해서는 월동기 평균온도 상승으로 기인할 것이라 여겨진다. 그러나 일조량 및 토양의 정밀 분석이 이루어지지 않았기 때문에 2년간 간장의 변화 원인에 관한 세밀한 연구가 필요하다.
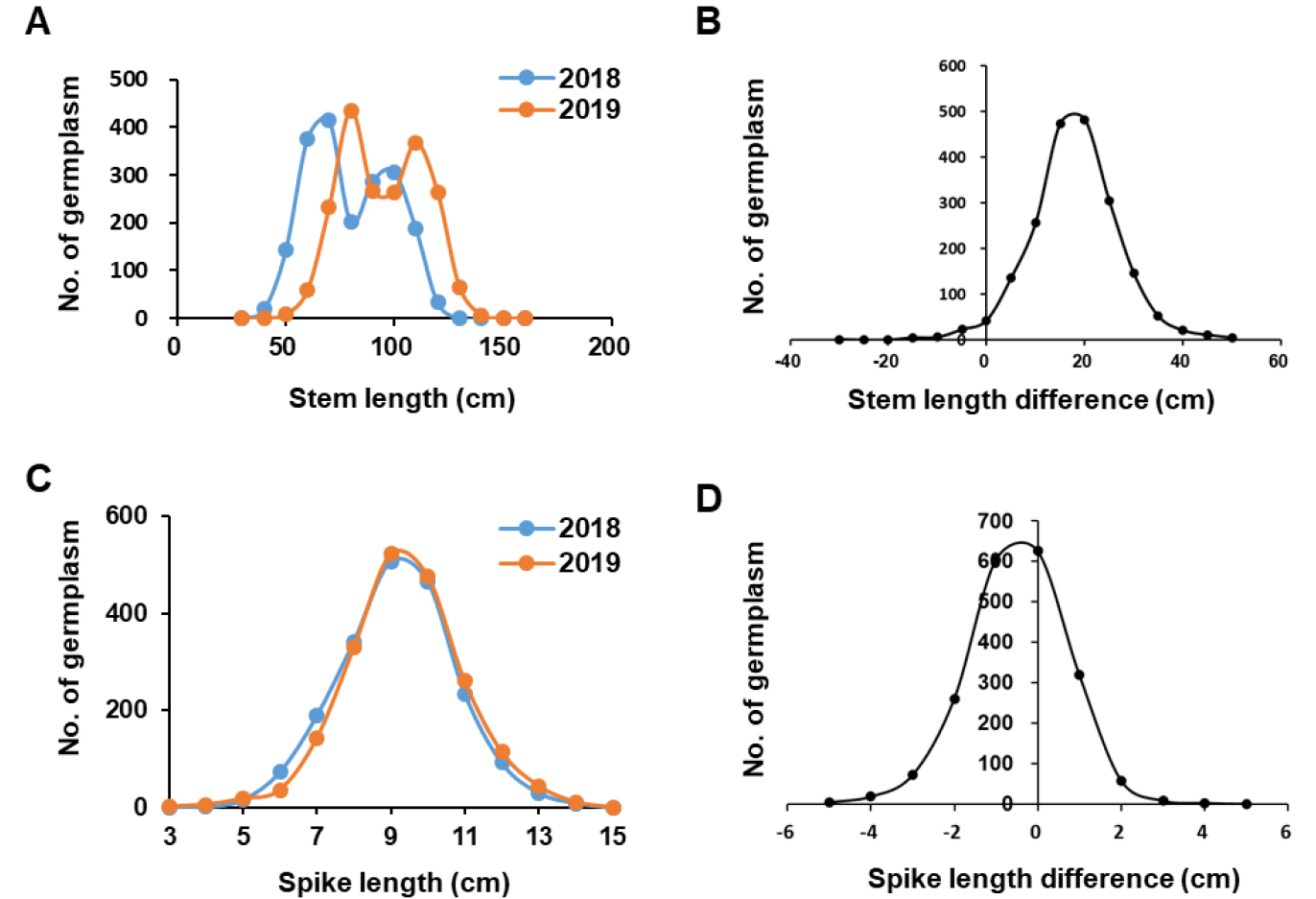
간장의 변화뿐만 아니라 도복된 밀 유전자원의 비율도 변화가 있었다. 2018년과 2019년 등숙기 도복율을 관능평가를 통해 온전히 존재하는 자원을 0으로 표기했으며, 완전히 쓰러진 자원을 9로 하여 0에서부터 9까지 6단계로 구분하여 3반복 평균값을 분석한 결과 2019년에 도복정도가 증가한 유전자원 수는 1826점으로써 도복 정도가 크게 증가하였다(Fig. 4A). 2018년도에 쓰러짐이 전혀 없는 자원(도복지수 0)의 수가 1,807점이였는데, 그 중에서 1,517점이 2019년에 완전히 도복되어 도복지수 9로 바뀌었으며 141점만이 그대로 도복지수 0을 유지하였다(Fig. 4A). 아울러 2018년과 2019년 모두에서 도복이 심한 9단계인 유전자원은 2점 이었다(data not shown). 또한 원산지별 도복율을 연차간 비교한 결과를 보면 한국 자원이 도복에 가장 강한 것을 확인할 수 있었다. 도복지수가 0인 자원의 비율이 전체 국내 자원의 25%를 차지하며 이 자원들은 2년간 도복없이 안정적인 생육을 보였다. 국내 자원 중 2018년 도복지수가 1이였던 유전자원이 2019년에 3으로 도복이 진행되었으며, 도복지수 4였던 74점과 도복지수 5였던 3자원이 2019년에는 모두 도복지수 9로 도복이 진행되었다. 아프가니스탄 자원의 80% 이상을 차지하는 도복지수 5인 자원과 그 이상의 도복지수를 보인 모든 자원이 도복지수 9로 모두 쓰러짐을 확인하였다. 중국자원의 경우도 2018년도에 도복지수가 5이상이던 85% 자원이 2019년에 모두 쓰러짐을 나타내었다. 이렇게 원산지에 상관없이 2018년에 절반정도 쓰러진 도복 현상을 보인 모든 자원들은 2019년에 가장 심각한 도복지수를 보였다.
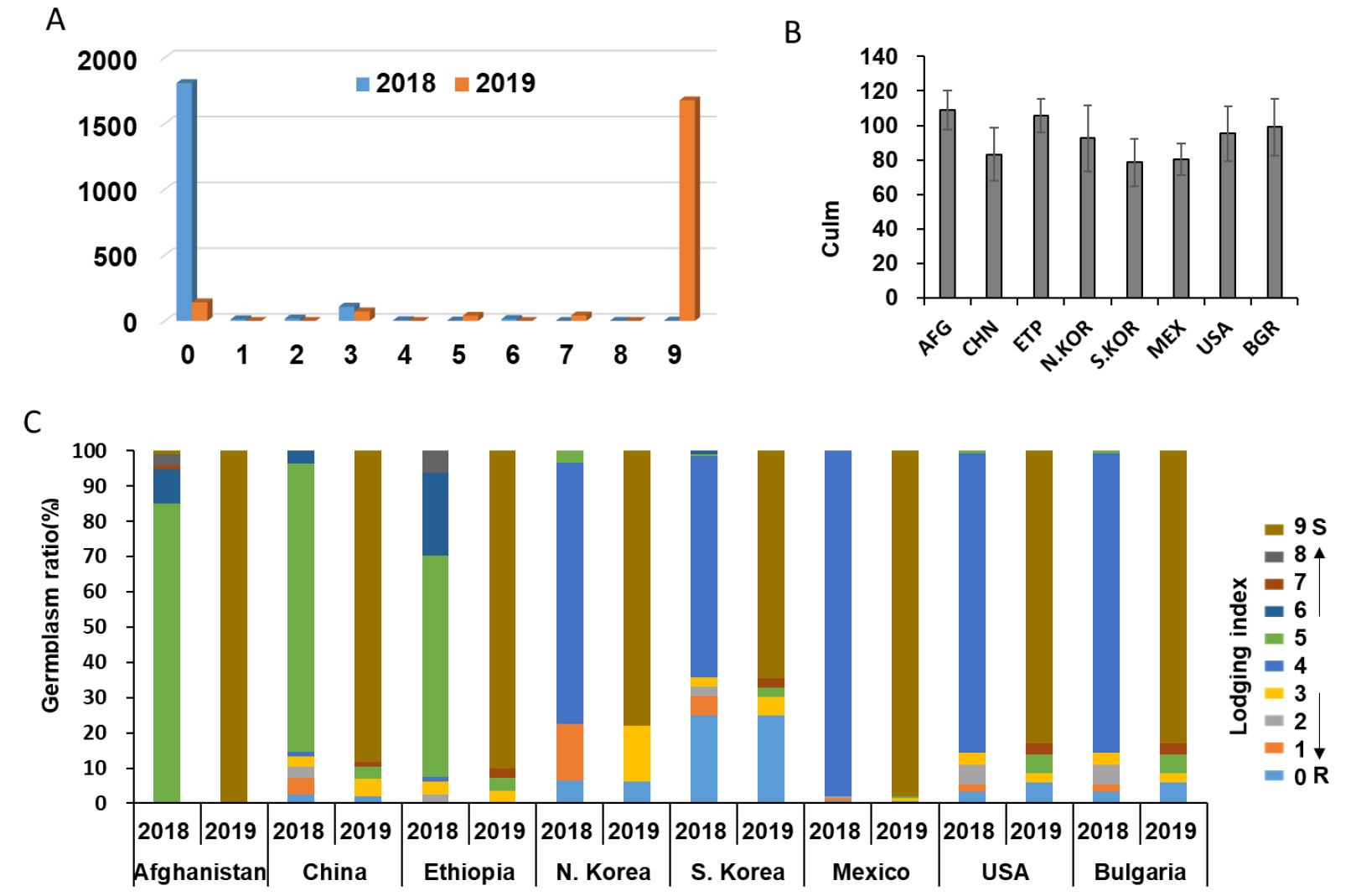
도복은 식물의 초장 및 절간 길이(Okuno et al, 2014), 줄기의 물리적 특성으로 줄기 속이 찬 정도 및 세포벽 두께 등(Xiang et al, 2016; Xiao et al, 2015; Pinera-Chavez et al, 2016), 줄기의 성분(lignin, cellulose 및 silicon) 함량 등(Okuno et al, 2014; Kong et al, 2013; Zhang et al, 2010), 관다발의 관다발 수 및 발달 정도(Hasnath & Jahan, 2013), 뿌리의 길이, 분포 면적, 굵기 등(Pinthus, 1967), 그리고 종실의 무게(Wang et al, 2006) 등이 관여한다. Okuno et al (2014)의 보고에 따르면 초장과 도복과의 연관은 없다고 보고하였으며, 본 연구의 초장과 도복과의 연관관계 분석에서도 연관이 없는 결과를 보였다(data not shown). 하지만, 2018년보다 2019년에 20 cm 가량 간장 길이가 길어진 것(Fig. 3A)과 2019년 도복 증가율이 증가한 것(Fig. 4A), 그리고 국가별 초장과 도복 정도(Fig. 4B, 4C)는 초장과 도복과의 연관성이 분명 존재하는 것으로 판단된다.
겨울철 기온 상승과 빠른 생육 재생기로 인한 영양생장 기간 변화와 간장의 증가와 도복 정도의 증가가 2018년 대비 2019년에 동시에 발생했지만, 그들간의 상관관계가 얼마나 존재하는지 여부는 밀 유전자원의 줄기의 특성 및 이삭의 무게 등 정밀한 분석을 통한 연구가 추가로 필요하다.
간장과 출수 및 성숙과의 관계
2년간 밀 유전자원의 간장과 출수와의 관계를 분석한 결과 상당한 상관관계가 존재함을 확인하였다(Fig. 5). 두 해 동안의 출수기와 간장과의 상관관계는 거의 유사하였으며(Fig. 5A, 5C), 간장은 또한 출수기와 마찬가지로 성숙기와도 상관관계를 보였다(Fig. 5B, 5D). 출수기와 성숙기 간의 상관관계는 높은 상관관계를 가지고 있기 때문에 간장과 동일한 정도의 상관관계를 나타났다(Supplementary Fig. 1). 그러나 많은 연구결과에서 밀의 초장과 출수기와의 상관관계가 존재하지 않는다고 보고하였다. Van Beuningen & Kokli (1990)는 얼룩무늬병 관련 유전자형 분석을 통해 밀의 초장과 출수기와의 상관관계 연구를 통해 밀의 초장과 출수기와의 상관관계가 없다고 보고하였다. Simon et al (2004)의 연구결과에서도 초장과 출수기와의 상관관계는 보이지 않았으나 곰팡이병 감수성이 출수기와 연관되었을 뿐, 초장과는 관계가 없다고 보고하였다. 많은 밀 육종가들은 출수기를 당겨 밀 생육기간을 단축시키고자 노력하였으며, 아울러 수량이 많은 밀을 선발함에 따라 증가될 수 밖에 없는 도복 피해를 줄이고자 초장이 작은 밀을 선발함으로써 밀 품종을 개발해 왔다. 그런 관점에서 본 연구결과에서 나타난 간장과 출수기와의 연관관계는 본 연구에서 사용된 수집 유전자원들이 대부분 육종가들에 목적에 따라 출수가 빠르며 도복에 강한 계통들이 대거 포함됨에 따른 현상으로 풀이된다(Khobra et al, 2019). 또한 초장과 출수기 간의 상관관계는 밀 생장점에서 분화된 유수가 성장하여 줄기 속을 통해 수가 지엽을 완전히 뚫고 나온 시점을 출수기로 판단하는 특성조사 방법으로 인해 초장이 긴 자원에서 수가 지엽 위로 출현하는데 까지 걸리는 시간 차이로 인해 발생하는 것으로도 여겨진다.
적 요
다양하고 우수한 밀의 유전자원을 확보하는 것은 앞으로 한국의 밀 육종에서 매우 중요한 임무이다. 따라서 본 연구는 한국뿐만 아니라 세계 60여개국으로부터 수집된 1967점의 유전자원을 확보하였고, 2018년부터 2019년까지 2년 동안 이들 자원에 대하여 특성 조사를 실시하였다. 최근 기후변화에 따라 고온과 이상기온 현상이 빈번하게 발생하고 있기 때문에, 한국의 환경에 적합하고, 특히 밀의 간장, 출수기, 성숙기 등 주요 농업 특성에 주목하였다. 2018년은 유수분얼기 및 출수기에 이상고온 현상이 나타났고, 2019년은 성숙기에 잦은 비로 인한 일조량 감소 현상이 있었다. 이와 같은 이유로 밀의 출수기와 성숙기의 변화가 확인이 되었다. 그러나 모든 자원이 같은 양상으로 변화하지는 않았다. 출수기와 성숙기가 빨라지거나 늦어졌고, 또한 기후에 상관없이 변화가 없는 밀도 있었다. 밀의 간장은 2019년이 2018년에 비해 평균 20 cm 증가하였다. 이 같은 이유는 2018년 월동기 기온의 상승이 원인일 것으로 생각되나 더욱 세밀한 연구가 필요하다. 또한 밀의 간장은 출수기와 성숙기와 상관관계를 보였고, 2018년과 2019년에 각각 35%와 20% 상관관계지수를 나타냈다. 이들 결과를 바탕으로 출수가 빠르고 성숙기간이 짧으며, 기후의 영향을 덜 받는 자원은 앞으로 한국의 밀 육종에 매우 유용하게 사용할 수 있을 것으로 생각된다.